|
|
| Un livre accompagne le site depuis octobre 2024. Découvrez-le ici > | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|


|
|
Connectome et neuromodulation : les deux grands systèmes du cerveau n’en forment qu’un
|
|
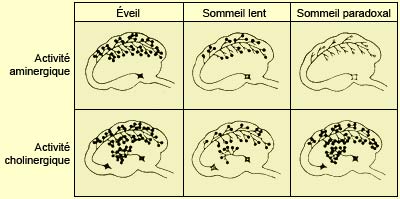
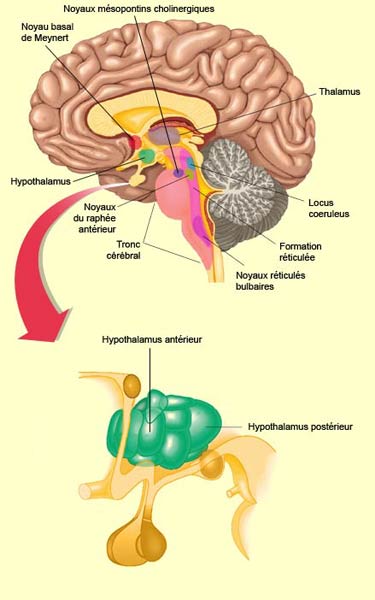
L’activation corticale nécessaire à l’éveil est rendue possible par un « réseau exécutif de l'éveil » comprenant l’hypothalamus postérieur, le thalamus intra-laminaires et le télencéphale basal. Mais ces structures sont elles-mêmes sous l’influence d’un réseau assez complexe d'une dizaine de structures situées en cascade du télencéphale basal au tronc cérébral inférieur et qui prennent en charge l'éveil. Très schématiquement, les composantes de ces systèmes modulateurs ascendants peuvent être groupées en deux grandes voies qui ont toutes deux leur origine dans une partie du noyau réticulé du bulbe rachidien. La voie ventrale se projette vers l’hypothalamus postérieur et le noyau de Meynert du télencéphale basal (neurones à acétylcholine) : c’est la voie réticulo-hypothalamo-corticale. La voie dorsale active les noyaux mésopontins cholinergiques, la formation réticulée mésencéphalique (neurones à asparte/glutamate) et le thalamus : c’est la voie réticulo-thalamo-corticale. En plus d’être à l’origine de l’éveil, plusieurs des noyaux de ces deux voies utilisant l’acétylcholine et le glutamate comme neurotransmetteur sont en partie responsables de l'activation corticale du sommeil paradoxal.
Parmi les noyaux les plus importants du tronc cérébral impliqués dans le système d’éveil, on retrouve donc :
L'ensemble de ces structures du tronc cérébral reçoit des collatérales des afférences sensorielles et végétatives qui participent ainsi au maintien de leur activité (voir encadré). Il s’agit d’un réseau complexe dont l'excitation pharmacologique d’un élément amène l'activation de tous les autres. Cette organisation redondante explique aussi pourquoi l'inactivation d'un seul système est suivie, après quelques jours, d’une récupération complète de l'éveil. Aucune des structures décrites, prise isolément, n'est donc indispensable à l'activation corticale.
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
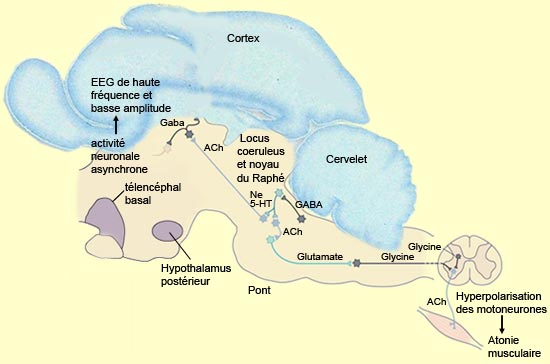
Le sommeil paradoxal, avec son EEG semblable à celui de l’éveil, est le produit de l’interaction complexe entre des noyaux du tronc cérébral, des structures du système limbique et des aires corticales. D’autres de ses caractéristiques étonnantes ont aussi des mécanismes qui ont pu être localisés. C’est le cas des mouvements oculaires rapides qui surviennent durant le sommeil paradoxal et dont on ignore la fonction. Ils sont produits par la formation réticulaire pontique et transmis aux couches motrices des colliculi supérieurs. Les neurones colliculaires projettent à leur tour sur la formation réticulaire pontique paramédiane (FRPP) qui coordonne la durée et la direction des mouvements oculaires. Autre caractéristique singulière du sommeil paradoxal dont on a localisé la source : la paralysie quasi-totale du corps qui l’accompagne. L’activité nerveuse intense enregistrée durant le sommeil paradoxal excite en effet la grande majorité des neurones corticaux, y compris ceux du cortex moteur primaire. Ces neurones moteurs génèrent ainsi des séquences d’activité organisées commandant des mouvements du corps mais seul les muscles respiratoires, oculaires, ainsi que ceux de l’oreille interne pourront actualiser la commande motrice. Celle-ci ne parviendra jamais jusqu’aux motoneurones des membres. Durant le sommeil paradoxal, l’augmentation d’activité de neurones cholinergiques de la protubérance va exciter d’autres neurones de la formation réticulée pontique qui utilisent le glutamate comme neurotransmetteur. Ceux-ci vont à leur tour envoyer des projections vers le bulbe rachidien où ils vont activer des interneurones qui relâchent de la glycine. Ce sont ces interneurones des noyaux réticulés bulbaires magnocellulaires, dont les axones descendent dans la moelle épinière, qui vont inhiber fortement les motoneurones en les hyperpolarisant.
Source: adapté de Appleton & Lange, Kandel, Schwartz, Jessell, Principles of Neural Science Quant aux pointes ponto-géniculo-occipitales (PGO) survenant de façon intermittente durant le sommeil paradoxal, elles seraient elles aussi issues de la formation réticulée pontique. Elles se propagent ensuite jusqu’au cortex occipital par l’entremise du thalamus bien qu’elles soient, comme leur nom l’indique, plus aisément détectables dans les relais du système visuel que sont les corps genouillés latéraux. Les ondes PGO font partie des différents événements
phasiques du sommeil paradoxal qui accompagnent les mouvements
oculaires rapides, les changements dans le rythme respiratoire
et cardiaque. Elles peuvent être générées
en l’absence de sommeil paradoxal en stimulant la protubérance
avec de l’acétylcholine, en particulier dans la
région péribrachiale. C’est dans cette région
réticulaire, autour du pédoncule cérébelleux
supérieur et sous le locus coeruleus, que sont générées
les ondes PGO. |
| |
|
|
|
|
|
|
|
|