Vouloir délimiter avec précision
les limites d’une région cérébrale particulière,
comme celle de Broca ou de Wernicke par exemple, pose de sérieuses difficultés.
On sait cependant que les aires cytoarchitectoniques décrites par Brodmann
représentent un meilleur corrélat anatomique que la forme des circonvolutions
cérébrales quand il s’agit d’établir des corrélats
anatomo-fonctionnels. Ceci dit, une aire corticale comme celle de Broca ne peut
être décrite de façon précise en se référant
uniquement aux aires de Brodmann. En effet, si pour plusieurs auteurs l’aire
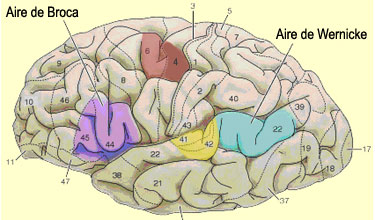
de Broca correspond aux régions 44 et 45 de Brodmann, pour d’autres
il s’agit tantôt uniquement de l’aire 44 ou 45, ou même
parfois de l’aire 44, 45 et 47.
Il n’est pas exclu non plus
que la partie la plus ventrale de l’aire 6 de Brodmann puisse également
en faire partie, de même que d’autres parties du cortex enfouies dans
les profondeurs du sillon latéral. L’hypothèse que certaines
sous-régions seulement de ces aires soient dévolues au langage ne
peut pas être exclu non plus.
L’acquisition du langage chez
l’être humain repose sur notre capacité d’abstraction
et d’utilisation de règles syntaxiques qui font défaut aux
autres animaux. On a par exemple montré lors d’expérience
en imagerie cérébrale qu’il y avait une activation de l’aire
de Broca lorsqu’un sujet apprenait une véritable règle de
grammaire d’une autre langue, mais pas quand cette règle décrivait
une opération linguistiquement illégale dans cette autre langue.
Il semble donc que des contraintes biologiques interagissent avec l’expérience
dans l’aire de Broca pour rendre possible l’acquisition d’une
langue.
L’aire de Broca devient ainsi un bon candidat de substrat
neuronal pour la « grammaire universelle » partagée par toutes
les langues du monde.
BROCA, WERNICKE ET LES AUTRES AIRES DU LANGAGE
On identifie généralement
l’aire
de Broca comme chevauchant les aires 44 et 45 de Brodmann situés à
l'avant de l'aire
prémotrice dans la région inféro-postérieur du
lobe frontal. Bien que ces régions contribuent toutes deux à notre
fluidité verbale, il semble que chacune ait une fonction particulière
et que l’aire de Broca puisse être ainsi séparée en
deux unités fonctionnelles.
D’une
part la partie postérieure du gyrus frontal inférieur
(aire 44) serait impliquée dans le traitement phonologique et la production
comme telle du langage, rôle qui serait facilité par sa position
proche des centres
moteurs de la bouche et de la langue. Et d’autre part la partie
antérieure de ce même gyrus (aire 45) qui serait davantage
impliquée dans les aspects sémantiques du langage. Sans être
directement impliquée dans l’accès au sens, l’aire de
Broca participe donc à la mémoire verbale (sélection et manipulation
d’éléments sémantiques).
Tout comme l’aire
de Broca, l’aire
de Wernicke n’est plus perçue comme une région anatomique
et fonctionnelle uniforme. L’analyse de plusieurs expériences en
imagerie cérébrale amène en effet à distinguer une
sous-région répondant à la parole (y compris celle du sujet)
et aux autres sons, une autre qui répond uniquement à des paroles
prononcées par quelqu’un d’autre en plus d’être
activée par le rappel d’une liste de mots, et une autre encore qui
serait davantage liée à la production de la parole qu’à
la perception. Ces résultats demeurent toutefois compatibles avec le rôle
général de cette région du lobe temporal gauche que l’on
associe à la représentation de séquences phonétiques,
qu’elles soient entendues, générées intérieurement
ou évoquées de mémoire par le sujet.
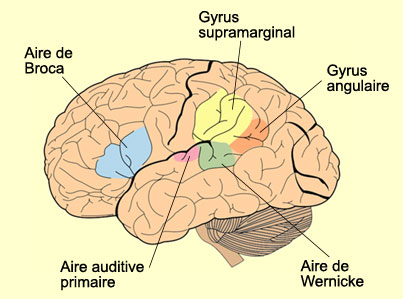
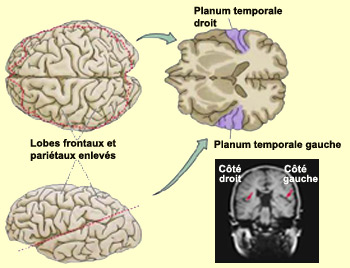
L’aire
de Wernicke, dont une composante anatomique clé est le
planum temporale, est située sur le gyrus temporal supérieur,
dans la portion supérieure de l’aire 22 de Brodmann. Il s’agit
d’une localisation stratégique compte tenu de ses fonctions dans
le langage. L’aire de Wernicke se trouve en effet située entre le
cortex auditif primaire (aires 41-42) et le lobule pariétal inférieur.
Ce dernier est principalement composé du deux régions
distinctes : caudalement, le gyrus angulaire (aire 39), lui-même
au voisinage des aires occipitales visuelles (aires 17, 18, 19), et dorsalement,
le gyrus supramarginal (aire 40) qui chevauche l’extrémité
du
sillon latéral, adjacent à la partie inférieure du cortex
somato-sensoriel.
Le
gyrus supramarginal semble impliqué dans le traitement
phonologique et articulatoire des mots tandis que le gyrus angulaire
serait impliqué davantage dans le traitement sémantique (de concert
avec le gyrus cingulaire postérieur). Le gyrus angulaire droit serait également
actif, révélant ainsi une contribution sémantique de l’hémisphère
droit dans le langage.
Les gyrus angulaire et supramarginal constituent
une région associative multimodale recevant des inputs
à la fois auditifs, visuels et somato-sensoriels. Les neurones de cette
région sont donc très bien placés pour traiter l’aspect
phonologique et sémantique du langage qui permet l’identification
et la catégorisation des objets.
Les aires du langage sont distinctes
des circuits de la perception auditive des mots entendus ou de la perception visuelle
des mots lus. Le cortex auditif permet la reconnaissance des sons, préalable
essentiel à la compréhension du langage. Le
cortex visuel, responsable pour sa part de la vision consciente du monde,
est aussi crucial pour le langage en permettant la lecture des mots et la reconnaissance
d’objets comme première étape de leur identification par un
nom.
Il existe une importante variabilité
de taille et de position des aires de Broca et de Wernicke selon les auteurs qui
les ont décrites.
Les
régions de haut niveau d’intégration sont plus hétérogènes
que les fonctions primaires, ce qui pourrait refléter une sensibilité
plus importante à l’influence de l’environnement et au phénomène
de plasticité.
L’organisation fonctionnelle du langage serait même variable chez
un même individu à différents moments de sa vie !
Une idée importante du modèle
de Mesulam est que la fonction d’une région dédiée
au langage n’est pas fixe mais varie selon le «contexte neural».
Autrement dit, la fonction d’une région particulière dépend
de la tâche à effectuer parce que ces régions n’activent
pas toujours les mêmes connexions entre elles. Ainsi, le gyrus frontal inférieur
gauche interagit avec des aires différentes selon qu’il traite le
son ou le sens d’un mot.
Ce type d’organisation en réseau
permet de sortir de l’équation « une aire = une fonction »
et explique beaucoup mieux des troubles
du langage parfois très spécifiques. Certaines personnes
sont par exemple incapables de nommer des outils ou de donner la couleur d’un
objet. D’autres encore peuvent expliquer la fonction d’un objet sans
pouvoir le nommer, et inversement.
Les études d’imagerie
cérébrale ont montré à quel point les tâches
cognitives comme celles impliquant le langage correspondent à un pattern
complexe d’activation de différentes aires distribuées sur
l’ensemble du cortex. Le fait qu’une aire cérébrale
particulière s’active lors de certaines tâches n’implique
donc pas qu’elle constitue à elle seule la localisation clairement
définie d’une fonction. En accord avec une conception plus distribuée
des fonctions cognitives qui prévaut de plus en plus en sciences cognitives,
cela signifie seulement que les neurones de cette région particulière
du cerveau sont davantage impliqués dans cette représentation que
leurs voisins. Cela n’exclut aucunement que d’autres neurones situés
ailleurs, et parfois très loin de cette région, puissent être
tout autant impliqués dans cette représentation.
Ainsi,
si le contenu d’un mot peut être encodé dans une assemblée
de neurones particulière, celle-ci n’est pas nécessairement
localisée à un seul endroit dans le cerveau. Au contraire, une assemblée
neuronale peut relier entre eux des neurones distribués dans des cortex
aux modalités distinctes (auditif, visuel, somatosensoriel ou moteur).
Car la compréhension ou la production d’une parole ou d’un
mot écrit nécessite l’apport concomitant de plusieurs de ces
modalités sensorielles. Les assemblées de neurones impliquées
dans le codage des fonctions grammaticales seraient pour leur part moins distribuées.
Il se pourrait donc que le cerveau traite les fonctions langagières
à la fois sur le mode parallèle par des réseaux distribués,
et à la fois grâce à des zones de convergence localisées.
MODÈLES CÉRÉBRAUX DU LANGAGE
PARLÉ ET ÉCRIT
Dans les années
1980, le neurologue Marsel Mesulam a proposé un modèle alternatif
à
celui de Wernicke-Geschwind pour appréhender les circuits du langage.
Il s’agit d’un modèle en réseaux hiérarchisés
où le traitement de l’information procède par paliers de complexité.
Par exemple, pour les traitements simples, comme dire les mois de l’année
dans l’ordre, les
aires motrices et prémotrices du langage sont directement activées.
Dire un énoncé nécessitant une analyse sémantique
et phonologique plus poussée fera pour sa part intervenir d’autres
aires en amont des aires motrices.
Pour ce qui est des paroles entendues,
elles sont perçues par l’aire auditive primaire, puis traitées
par des aires corticales dites associatives unimodales: les régions temporales
supérieures et antérieures ainsi que la région operculaire
du gyrus frontal inférieur gauche.
Selon le modèle de Mesulam,
ces aires unimodales transmettent ensuite leurs informations à deux sites
d’intégration distincts : au pôle temporal appartenant au système
paralimbique donnant accès au système de mémoire
à long terme et au système émotionnel;
et à la partie terminale postérieure du sillon temporal supérieur
qui permet l’accès au sens. Les régions triangulaire et orbitaire
du gyrus frontal inférieur participent aussi au traitement sémantique.
Localisation
approximative du gyrus frontal inférieur. Il est divisé en trois
parties : operculaire, triangulaire et orbitale. La partie triangulaire du gyrus
frontal inférieur forme l’aire de Broca.
Mesulam
considère qu’il existe néanmoins deux « épicentres
» du traitement sémantique, soit les
aires de Broca et de Wernicke. Cette nouvelle conception des aires de Broca
et de Wercnicke concorde avec le fait que ces deux aires travaillent souvent de
façon synchrone lors de la réalisation d’une tâche de
traitement des mots, appuyant l’existence de connexions très fortes
entre elles.
Ce concept d’épicentre rejoint celui de zones
de convergences proposé par d’autres auteurs. Il s’agit
de zones où l’information en provenance de différentes modalités
sensorielles peut être mise en commun.
Cette mise en commun se
ferait par la formation d’assemblées cellulaires, c’est-à-dire
un groupe de neurones interconnectés dont les synapses ont été
renforcées suite à leur activation simultanée (la «règle
de Hebb» ). Cette conception des aires du langage comme des zones de
convergence où s’établissent des assemblées neuronales
accorde donc une place importante à l’influence épigénétique
dans l’apprentissage d’une langue.
L’une de ces zones
de convergence est sans contredit le lobule pariétal inférieur gauche
qui comprend le gyrus angulaire et le gyrus supramarginal.
En plus de recevoir de l’information de l’hémisphère
droit, le lobule pariétal inférieur gauche intègre aussi
des associations émotionnelles en provenance de l’amygdale
ou du gyrus cingulaire.
Certains croient d’ailleurs que le langage
est demeuré sous contrôle limbique
jusqu’à ce que le lobule pariétal inférieur évolue
et deviennent ce lieu de convergence riche en inputs pour l’aire
de Broca. D’aucuns pensent aussi que c’est l’émergence
du lobule pariétal inférieur qui a rendu l’être humain
capable de segmenter les sons entendus pour qu’ils fassent sens et inversement
d’exprimer de manière séquentielle des sons qui signifient
des choses. C’est ainsi que les vocalisations émotionnelles et sociales
primitives en seraient venues à être gouvernées par des règles
grammaticales d’organisation pour produire ce que l’on appelle le
langage moderne.
Enfin, plusieurs chercheurs
rejettent aujourd’hui les modèles localisationnistes classique comme
ceux de Geschwind
ou de Mesulam. À la place, ils proposent une conception du langage et des
fonctions cognitives en général comme étant distribuées
sur des aires anatomiquement distinctes qui traitent l’information en parallèle
(plutôt que de façon sérielle d’une «aire du langage»
à une autre).
Les tenants de cette vision «en parallèle»
du traitement de l’information linguistique acceptent tout de même
une certaine localisation des fonctions primaires, tant auditives qu’articulatoires.
Ce concept d’un réseau de traitement parallèle et
distribué de l’information linguistique constitue un paradigme
épistémologique distinct qui inspire la réévaluation
de certaines études d’imagerie cérébrale fonctionnelle.
Pour les tenants de ce paradigme, l’étendue de l’activation
des différentes zones de l’hémisphère gauche ainsi
que le grand nombre des processus psychologiques impliqués exclut une association
précise des fonctions à des aires anatomiques. Seulement pour le
rappel des mots par exemple, il est le produit d’un réseau très
distribué localisé de façon prédominante à
gauche et incluant le lobe temporal inférolatéral, le lobe pariétal
postérieur inférieur, les régions prémotrices du lobe
frontal, le gyrus cingulaire antérieur et l’aire motrice supplémentaire.
Quant à la contribution de chaque région à cette tâche
spécifique, un réseau aussi largement distribué exclut, pour
les tenants du traitement en parallèle, l’attribution précise
de fonctions à des structures.
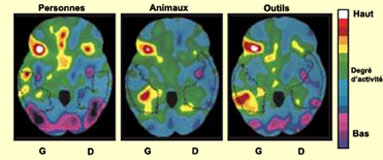
L’accès au sens semble
fonctionner selon des catégories qui sont localisées physiquement
dans le cerveau : la catégorie « personnages célèbres
» est perdue quand le pôle temporal (l'extrémité antérieure
du lobe temporal) est touché et la catégorie « animaux »
quand la lésion a lieu dans la partie intermédiaire et inférieure
du lobe temporal. Il semble aussi que les réseaux qui participent à
l’encodage des mots fassent appel à des aires faisant partie du système
moteur ou visuel. Nommer des outils active les aires prémotrices frontales
alors que nommer des animaux recrute des aires visuelles, alors que dans les deux
cas les aires de Broca et de Wernicke ne sont même pas activées.
Parmi les chercheurs qui plaident
en faveur d’un système de traitement du langage distribué
à travers différentes structures cérébrales, certains
comme Philip Liberman accordent une grande importance au rôle que joueraient
les
ganglions de la base dans le langage. D’autres structures sous-corticales
traditionnellement impliquées dans le contrôle moteur, comme le
cervelet ou le thalamus, seraient également impliquées. Ces
conceptions s’opposent à celles de Chomsky sur la nature exceptionnelle
du langage humain pour se situer résolument dans une
perspective évolutive adaptationniste.
Chez de nombreuses espèces
aussi loin de nous que la grenouille, le cerveau est latéralisé
à gauche pour la fonction de vocalisation.
Pour les régions
anatomiques qui correspondent aux aires de Broca et de Wernicke, la latéralisation
cérébrale existe déjà chez le chimpanzé même
si elle ne correspond pas encore à la fonction du langage. Et comme chez
l’être humain, la majorité des chimpanzés utilisent
préférentiellement leur main droite plutôt que la gauche.
Ces asymétries chez les autres primates constituent autant de
preuves de l’origine phylogénétique ancienne de la latéralisation
cérébrale. L’expansion
massive du cortex préfrontal pourrait d’ailleurs en partie
refléter son rôle dans la production du langage.
Les femmes ont la réputation
de pouvoir parler et écouter en faisant toutes sortes de choses en même
temps, alors que les hommes préféreraient parler d’une chose
ou en écouter une autre, mais successivement plutôt que simultanément.
L’imagerie cérébrale a peut-être révélé
un fondement anatomique à cette différence de comportement en montrant
que le langage sollicite davantage les deux hémisphères chez les
femmes et serait plus latéralisé (surtout à gauche) chez
les hommes. Davantage de fibres nerveuses relient aussi les deux hémisphères
cérébraux chez la femme, ce qui va aussi dans le sens de plus grands
échanges d’information entre les deux hémisphères.
PRÉFÉRENCE MANUELLE, LANGAGE ET
LATÉRALISATION CÉRÉBRALE
Il est clair que l’asymétrie
anatomique du cerveau, la
latéralisation pour le langage et la préférence manuelle
sont liés mais cette influence mutuelle est complexe. Bien qu’environ
90 % des gens sont droitiers, et qu’environ 95% de ceux-ci ont leurs aires
du langage à gauche, il n’en demeure pas moins que certains droitiers
ont une latéralisation pour le langage à droite ou répartie
dans les deux hémisphères. Sans parler des gauchers où
l’on retrouve tous les cas de figure, y compris la latéralisation
à gauche.
Certains suggèrent d’ailleurs que la dominance
de l’hémisphère gauche pour le langage a évolué
à partir du meilleur contrôle de cet hémisphère pour
la main droite. Les circuits contrôlant la main habile pourraient avoir
évolué pour prendre en charge les circuits moteurs impliqués
dans le langage. L’aire
de Broca, en particulier, est fondamentalement un module prémoteur
du néocortex et coordonne des patterns de contractions musculaires qui
ne sont pas seulement reliés au langage.
Les
études d’imagerie cérébrale ont mis en évidence
plusieurs structures cérébrales impliquées dans le langage
qui s’avèrent plus grandes dans l’hémisphère
gauche que dans le droit. Bien que l’aire de Broca située dans le
lobe frontal gauche est plus grande que son homologue dans l’hémisphère
droit, les asymétries les plus fortes se retrouvent surtout dans les aires
postérieures du langage comme le planum
temporale ou le gyrus angulaire par exemple.
Une autre asymétrie notable
est celle de l’extension du lobe frontal droit et du lobe occipital gauche.
Ces extensions pourraient cependant provenir d’une légère
rotation des hémisphères (dans le sens anti-horaire lorsque vus
de haut) plutôt que d’une différence de volume de ces régions.
On appelle ces asymétries les «petalias» frontales et occipitales,
un terme originellement employé pour désigner l’indentation
fait à l’intérieur du crâne par ces extensions.
Les structures impliquées dans la production et la compréhension
du langage semblent se mettre en place selon des commandes génétiques
qui entrent en jeu dès la
migration neuronale. Cela n’empêche pas que les deux hémisphères
puissent demeurer à peu près équipotent jusqu’à
l’acquisition
du langage. Normalement, la spécialisation langagière bascule
vers l’hémisphère gauche dont la maturation serait légèrement
plus hâtive. L’activité neuronale plus précoce et plus
intense de l’hémisphère gauche mènerait alors à
l’usage préférentiel de la main droite et à la prise
en charge des fonctions langagières.
Mais si l’hémisphère
gauche est lésé ou déficient, le
langage peut être acquis par l’hémisphère droit.
Une surabondance de testostérone fœtale due à un stress à
la naissance pourrait bien être l’une des causes les plus fréquentes
de ralentissement du développement de hémisphère gauche entraînant
une plus grande participation de l’hémisphère droit.
Cette hypothèse d’un rôle central de la testostérone
est appuyée par des expériences faites sur des rats et qui ont montré
que l’asymétrie corticale est modifiée si l’on injecte
aux rongeurs de la testostérone à la naissance. L’hypothèse
hormonale expliquerait également pourquoi les deux tiers des gauchers sont
des hommes.
Les variations interindividuelles,
indispensables pour la sélection naturelle, s’expriment de différentes
façons dans le cerveau humain. Le volume et le poids peuvent varier d’un
facteur de deux et même plus. L’organisation vasculaire est extrêmement
variable, une obstruction à un point donné du système vasculaire
pouvant amener des déficits fort différents d’un individu
à l’autre. Au niveau anatomique macroscopique, les circonvolutions
et les scissures corticales varient aussi énormément d’un
individu à l’autre, en particulier dans les
aires associées au langage. Une variabilité s’observe
également au niveau microscopique, par exemple dans la variabilité
de la structure synaptique des neurones de l’aire de Wernicke.
Enfin, une variabilité interindividuelle s’exprime dans l’organisation
fonctionnelle du cerveau, en particulier dans le phénomène d’asymétrie
hémisphérique. Des données indiquent par exemple que les
fonctions langagières pourraient être davantage bilatérales
chez la femme que chez l’homme. Le pourcentage de latéralisation
atypique pour le langage diffère également selon la préférence
manuelle, étant considérablement plus élevée chez
les gauchers que chez les droitiers.
Enfin, comme si ce n’était
pas déjà assez, on peut aussi parler d’une variabilité
« intra-individuelle », une fonction cérébrale donnée
pouvant parfois solliciter des assemblées neuronales distinctes selon différentes
circonstances, une nouvelle tâche versus cette même tâche une
fois entraîné, par exemple.
Plusieurs hypothèses tentent
d’expliquer l’origine de la pragmatique, c'est-à-dire de notre
capacité à prendre en compte le contexte interpersonnel dans notre
utilisation du langage. C’est le cas de la
théorie de l’esprit. Celle-ci consiste selon Premack
et Woodruff (1978) en l’habileté permettant aux gens d’attribuer
des phénomènes mentaux à autrui, de raisonner à partir
de ceux-ci et de comprendre les comportements qui en découlent. Ces auteurs
ont été les premiers à utiliser le terme « théorie
de l’esprit » et ce, dans le cadre d’une étude sur la
capacité des chimpanzés à inférer des croyances et
des intentions à un humain. Cette théorie a depuis surtout été
étudiée dans une perspective
développementale chez des enfants normaux et autistes, ces derniers
formant une population désormais reconnue pour présenter, et ce
dès les premières étapes du développement, des déficits
en matière de raisonnement social.
Si l’on demande à des
individus d’identifier la teneur émotive de phrases enregistrées
qu’on leur fait entendre dans une seule oreille, ils performent mieux lorsque
c’est l’oreille gauche (qui envoie l’information à l’hémisphère
droit) que lorsque c’est l’oreille droite (qui l’envoie à
l’hémisphère gauche).
CONTRIBUTION DE L'HÉMISPHÈRE DROIT
AU LANGAGE
Pour suivre une conversation,
comprendre un texte ou une plaisanterie, on doit non seulement être capable
de comprendre la syntaxe des phrases et le sens des mots mais également
de mettre en relation plusieurs éléments et de les interpréter
par rapport à un contexte donné. Si des lésions dans l’hémisphère
gauche produisent les
nombreux types d’aphasies connus, celles qui surviennent dans l’hémisphère
droit provoquent une variété de déficits que l’on peut
regrouper en deux grandes catégories.
Certains exercent d’abord
un effet indirect sur la communication en perturbant l’habileté à
interagir de façon compétente avec son environnement.
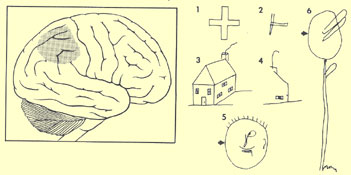
La personne cérébrolésée
du côté droit peut par exemple souffrir d’héminégligence,
un déficit qui fait en sorte que le patient néglige ou ignore les
stimuli de différentes modalités sensorielles présentés
du côté controlatéral à la lésion.
Les
dessins 2, 4, 5 et 6 ont été faits par un patient héminégligent.
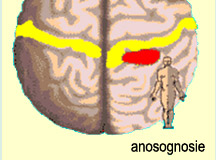
Elle peut aussi être atteinte
d’anosognosie,
c’est-à-dire une non-reconnaissance de ses déficits. En fait,
il arrive même aux personnes qui ont une lésion juste à l’arrière
de la scissure centrale droite de ne pas reconnaître certaines parties de
son propre corps comme étant les siennes. On voit donc que ces lésions
à l'hémisphère droit produisent une sorte d’indifférence
qui est à l'opposé de l'investissement émotionnel minimal
nécessaire à l’établissement d’une communication
harmonieuse.
L’autre grande famille de déficits
que l’on peut observer suite à une lésion dans l’hémisphère
droit affecte directement la communication et la cognition. Ils peuvent être
regroupés sous le vocable de troubles pragmatiques, la pragmatique
étant cette discipline qui s’intéresse aux rapports entre
le langage et l’utilisation contextuelle qu’en fait ses utilisateurs.
Parmi les troubles pragmatiques, on distingue ceux de la prosodie, ceux de l’organisation
du discours et ceux de la compréhension du langage non-littéral.
Activité
du cerveau d’une femme qui doit décider si des mots riment ou non.
On voit que l’hémisphère droit est grandement sollicité.
Source:
Shaywitz & Shaywitz, Yale Medical School.
La
prosodie fait référence à l’intonation et
à l’accentuation des phonèmes. Ainsi, un patient à
l’hémisphère droit cérébrolésé
dont la prosodie est déficiente est incapable d’exprimer une émotion
réellement ressentie de façon adéquate. Par conséquent
la personne souffrant d’aprosodie se comporte et tient des propos semblant
manquer d’affectivité.
Autre aspect de la pragmatique pouvant
être touché par des lésions à l’hémisphère
droit est le discours ou plutôt son organisation
découlant des règles qui régissent sa construction. On note
ainsi chez certains patients une moins grande capacité à distinguer
les indices permettant d’établir un contexte communicationnel, les
nuances apportées par certains mots, les intentions du locuteur, le langage
corporel ou les conventions sociales. Concernant les conventions sociales par
exemple, on n’aborde généralement pas son frère ou
son patron de la même façon, distinction que font pourtant difficilement
certains cérébrolésés droits.
Enfin,
dernière catégorie de trouble pragmatique mais non la moindre, la
compréhension du langage non-littéral. On estime en effet
que plus de la moitié des phrases que l’on prononce ne désigne
pas littéralement ce qu’on veut dire, du moins pas totalement. C’est
le cas de l’ironie, des métaphores et des actes de langage indirects,
tous reliés aux intentions des locuteurs.
L’ironie par
exemple, qui constitue un état d’esprit de second niveau au même
titre que les mensonges et les blagues, implique de saisir l’état
d’esprit d’un interlocuteur (premier niveau), de même que ses
intentions concernant la façon dont ses dires devraient être perçus
par autrui (second niveau). On peut alors comprendre que le blagueur souhaite
ne pas être pris au sérieux, alors que l’ironique s’attend
à ce que le contraire de son propos soit perçu comme le message
final.
Les métaphores traduisent également une intention
qui n’est pas conforme avec l’interprétation littérale
du propos. Ainsi, si un élève souffle à son voisin de classe:
« Ce professeur est un somnifère. », le voisin comprend l’analogie
sous-entendue entre la pilule et le professeur et conclue que l’élève
trouve le professeur endormant, ce qui ne serait pas nécessairement le
cas d’un patient cérébrolésé droit.
Finalement, les actes de langage indirects, couramment utilisés dans la
vie quotidienne, peuvent aussi causer quelques soucis aux cérébrolésés
de l’hémisphère droit. Ici, l’intentionnalité
est sous-jacente au message verbal énoncé en tant que tel. Par exemple,
lorsque quelqu’un mentionne : « Je ne sais pas quelle heure il est.
», on comprend tout de suite que la personne demande indirectement si quelqu’un
peut lui dire l’heure. Ce qui n’est pas le cas des cérébrolésés
droits.
Même si l’hémisphère
gauche est encore vu comme l’hémisphère dominant en matière
de langage, le rôle de l’hémisphère droit dans la prise
en compte du contexte est donc maintenant établi. Et si en l’absence
de l’hémisphère gauche (par exemple lors d’un test
de Wada), le droit peut produire un langage rudimentaire, à la lumière
des études lésionnelles son rôle dans le langage apparaît
beaucoup plus vaste. De sorte qu’il est plus juste d’envisager les
spécialités langagières des deux hémisphères
non plus comme des fonctions séparées, mais bien comme diverses
habiletés fonctionnant en parallèle et dont l'interaction rend possible
le langage humain dans toute sa complexité.