Un livre accompagne le site depuis octobre 2024. Découvrez-le ici >
De l'embryon à la morale
Un autre processus qui contribue
à mener les axones à bon port est celui de la fasciculation
par lequel les neurones qui se développent ensemble s’associent en
se chevauchant les uns sur les autres. Comme la surface de la matrice extracellulaire
est limitée, cela permet à plusieurs neurones de s’empiler
littéralement sur celui qui avance sur le substrat. La fasciculation est
rendue possible par l’expression de molécules présentes sur
la surface des membranes neuronales appelées molécules
d’adhésion (ou CAM pour Cell
Adhesion Molecules, en anglais). Les CAMs des axones voisins interagissent donc
pour permettre l’adhésion mutuelle de ceux-ci et une croissance groupée
vers leur cible.
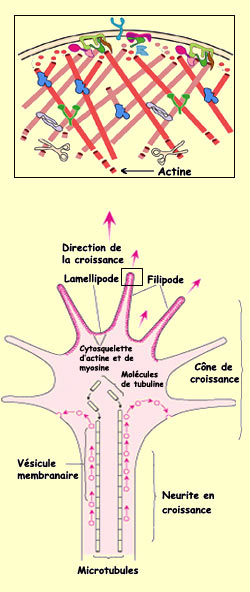
LE CÔNE DE CROISSANCE
Le cône de croissance
est la structure amiboïde située au bout de l’axone en croissance
et qui le tire vers l’avant. La motilité du cône de croissance
est ponctuée de phases de protrusion, d’adhésion et de contraction.
La protrusion du cône de croissance dans la bonne direction dépend
de toute une machinerie cellulaire qui répond aux signaux de l’environnement
par l’entremise de récepteurs spécialisés. On sait
par exemple que c’est la polimérisation de l’actine
qui provoque l’avancement du cône de croissance et que la cascade
biochimique qui déclenche cette polimérisation implique des GTPases
de la famille Rho.
Les filaments d’actine dans l’axone en
croissance se projettent dans les filipodes. De manière moins organisée,
ils envahissent aussi les lamellipodes pour former une sorte de treillis. Près
de la périphérie du cône de croissance, les filaments d’actine
s’allongent par polymérisation des protéines d’actine.
À l’opposé, vers la région centrale des lamellipodes,
les filaments d’actine subissent un désassemblage. C’est d’ailleurs
ce même processus que l’on retrouve chez toutes les cellules qui se
déplacent par protrusion de lamellipodes.
Au centre de l’axone
qui s’allonge derrière le cône de croissance en progression,
un cytosquelette fait de protéines de tubuline se forme.
Des vésicules voyagent le long de ces microtubules et viennent fusionner
avec la membrane du cône de croissance pour lui permettre de prendre de
l’expansion.
Source : Max-Planck-Institut
La croissance de l’axone dépend
de l’interaction moléculaire entre le cône de croissance et
son substrat. L’une des composantes principales de ce substrat est la matrice
extracellulaire formée des protéines fibreuses situées entre
les cellules. À travers cette matrice se dessine de véritables corridors
balisés par des protéines permissives comme la laminine
par exemple. Celle-ci se lie avec des molécules appeléesintégrines situées sur la membrane du cône
de croissance favorisant la progression du cône de croissance dans la bonne
direction. D’autres protéines, répulsives celles-là,
se trouvent également sur le substrat pour aiguiller encore davantage l’élongation
de l’axone en l’empêchant de prendre certaines directions. L’élongation
des axones est ainsi soumise à une véritable signalisation routière
le long « d’autoroutes moléculaires ».
Un des grands défis pour
les neurones de nos systèmes moteurs et sensoriels est de faire en sorte
que des points adjacents dans la partie périphérique de ces systèmes
soient représentés également par des points contigus dans
les centres de traitement cérébraux. Comment les axones en croissance
font-ils pour s’organiser avec une telle précision pour former des
cartes topographiques aussi précises que l’homonculus
moteur ou somatosensoriel, par exemple ?
Suite à des expériences
sur les voies visuelles de la grenouille au début des années 1960,
on a émis l’hypothèse d’une chimioaffinité entre
des molécules sur le cône de croissance des différents axones
et d’autres sur les cellules cibles où les ces axones doivent aboutir.
Un peu comme si chaque axone en provenance de la périphérie recevait
une étiquette chimique indiquant sa position et que des étiquettes
complémentaires sur les cellules cibles leur permettaient de se situer
selon cette même position.
Mais cette hypothèse d’une
affinité de type «clé-serrure» où il y aurait
autant de type de clés et de serrures différentes qu’il y
a de positions à coder s’est cependant révélée
contredite par les données expérimentales. En fait, d’autres
expériences ont montré qu’il y a bien une correspondance entre
des molécules du cône de croissance et d’autre de la structure
cible, mais cette correspondance se fait plutôt grâce à un
gradient de concentration qui n’utilise qu’un seul jeu de clés
et de serrure.
Source: Dr. Brian E. Staveley Department
of Biology Memorial University of Newfoundland
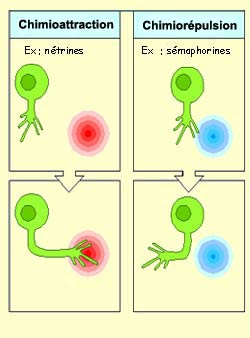
La
famille de molécules diffusibles chimioattractives la mieux caractérisée
est celle des nétrines. Ces molécules sécrétés
par des cellules cibles diffusent dans le milieu extracellulaire et influencent
à distance le cône de croissance de certains neurones. Les nétrines
ressemblent aux molécules d’adhérence cellulaire situées
dans la matrice extracellulaire comme la laminine qui guident le cône de
croissance par contact direct. Comme pour ces dernières, les nétrines
influencent le cône de croissance en se fixant sur des récepteurs
transmembranaires spécifiques qui vont assurer la transduction du signal.
Dans l’embryon, la sécrétion des nétrines
se fait souvent près de la ligne médiane où les axones ont
à choisir s’ils restent du même côté ou s’ils
traversent cette ligne. On a bien décrit son rôle par exemple dans
le développement du faisceau spinothalamique qui relie les informations
thermiques et nociceptives de la périphérie jusqu’au thalamus.
Les nétrines aident aussi des axones du système nerveux central
à franchir la ligne médiane, comme ceux du corps calleux ou du chiasma
optique par exemple.
Du côté de la chimiorépulsion,
on retrouve la famille des sémaphorines, des molécules
qui repoussent les cônes de croissance des axones. Ces molécules
peuvent être soit fixées à la membrane cellulaire, soit sécrétées
et diffusées dans le milieu environnant.
Les
molécules de guidage de la croissance axonale ne sont pas attractives ou
répulsives en soi. Elles acquièrent plutôt l’une des
deux caractéristiques selon le type de récepteur exprimé
sur un cône de croissance particulier. Car comme pour la transmission synaptique
et de nombreux autres processus à l’origine de cascades de réactions
biochimiques, c’est l’affinité électrochimique entre
une molécule et son « récepteur » qui met en branle
les mécanismes internes qui vont amener le cône de croissance à
changer de direction.
Ainsi, la molécule
de guidage bien connue nétrine-1 est attractive pour les
interneurones de la commissure dorsale mais agit comme un facteur répulsif
pour certaines classes de motoneurones. Un facteur de guidage donné peut
donc être interprété de deux façons complètement
différentes par le cône de croissance selon le type de récepteurs
qu’exprime ce cône à sa surface.
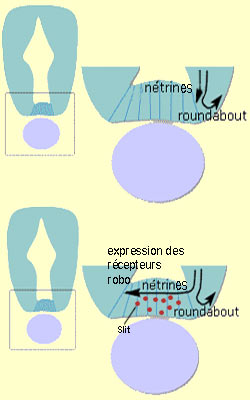
Pour poursuivre avec
notre exemple, attiré dans la région médiane de la partie
ventrale de la
moelle épinière par les nétrines, les cônes de
croissances des interneurones se mettent alors à exprimer davantage le
récepteur roundabout (ou robo) sensible
à un facteur de répulsion appelé slit lui
aussi sécrété dans la région médiane ventrale
de la moelle. Par conséquent, les cônes de croissance se mettent
alors à fuir cette région ce qui les amène à poursuivre
de l’autre côté de la ligne médiane.
Section du tube neural avec un agrandissement de la
région du plancher (l’ovale mauve représente la notochorde).
Dans cet exemple, les cellules de la ligne
médiane de la moelle épinière qui sécrètent
ces différentes molécules de guidage agissent comme des cibles intermédiaires
qui dans un premier temps attirent l’axone en croissance, et dans un second
temps le repousse vers sa cible définitive.
Le NGF est produit par les cellules
cibles du système
nerveux sympathique. Si on le neutralise avec l’injection d’anticorps
spécifiques, les ganglions sympathiques dégénèrent
complètement. Même chose si l’on interrompt le transport axoplasmique
: les neurones meurent malgré la libération de NGF par le tissu
cible.
Le NGF est une
protéine qui contient 3 sous-unités, alpha, beta et gamma. La sous-unité
gamma a des propriétés enzymatiques protéolytiques. La sous-unité
alpha inhibe cette protéase. La sous-unité beta est responsable
de l’activité biologique.
Cette protéine
ressemble à d’autres protéines favorisant la croissance (comme
l’insuline), ce qui suggère qu’elles proviennent de la même
famille de molécule. Il existe de nombreux autres facteurs de croissance
neuronale, les motoneurones n’étant par exemple pas sensibles au
NGF.
FACTEURS DE CROISSANCE ET MORT NEURONALE
Au cours d’une
longue période qui commence avant la naissance et dure jusqu’à
l’adolescence, on assiste à une réduction très importante
du nombre de neurones et de synapses nouvellement élaborées. Cette
mort neuronale découle d’une compétition pour des facteurs
trophiques émis en quantité limitée par les cellules cibles.
Le premier de ces facteurs à avoir été mis en évidence
au début des années 1950 (par la biologiste italienne Rita Levi-Montalcini,
Prix Nobel avec Stanley Cohen en 1986) est un peptide connu sous le nom de facteur
de croissance des nerfs (NGF pour Nerve Growth Factor,
en anglais). Le NGF est reconnu par les axones sympathiques et transporté
de façon rétrograde à l’intérieur de l’axone
jusqu’au corps cellulaire où ses interactions moléculaires
favorisent la survie du neurone.
En fait, le NGF et les
autres neurotrophines procèdent plutôt en verrouillant un programme
génétique d’autodestruction que possède chaque cellule.
Cette destruction programmée du neurone a pour nom l’apoptose
(voir capsule outil à gauche). Contrairement à la nécrose
qui est un processus de mort accidentelle de la cellule et qui provoque de l’inflammation,
l’apoptose est une déstructuration méthodique des composantes
de la cellule qui se fait en douceur. Ce processus de mort cellulaire programmée
permet une configuration optimale du câblage cérébral en sélectionnant
les cellules dont les connexions s’avèrent les plus efficaces, en
équilibrant la quantité de neurones pré et postsynaptiques
ainsi qu’en éliminant les neurones malformés ou endommagés.
Les différentes neurotrophines n’agissent pas toutes sur les
mêmes neurones. L’action du NGF permet par exemple la survie des neurones
parasympathiques, celle du BDNF de neurones appartenant à des ganglions
sensitifs, et celle de la NT-3 à la survie de ces deux populations. Cette
sélectivité d’action des neurotrophines a son origine dans
une famille de récepteurs appelés Trk, désignée ainsi
parce que ces protéines ont été initialement identifiées
comme des récepteurs capables d’activer l’enzyme tyrosine kinase.
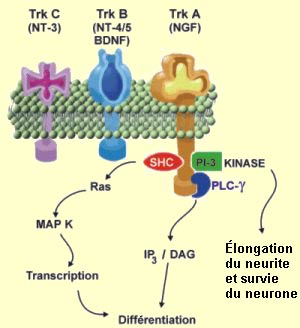
Source : Dr. Paul Kelly
Dans
la famille des récepteurs Trk aux neurotrophines, on retrouve
le TrkA qui est principalement un récepteur du NGF, le TrkB qui est un
récepteur du BDNF et le TrkC qui est un récepteur de la NT-3. Étant
donné la grande ressemblance de structure entre les neurotrophines d’une
part, et entre les récepteurs Trk d’autre part, il existe un certain
degré d’activation croisée entre eux. Ainsi, dans certaines
conditions, la NT-3 peut se lier au récepteur TrkB et l’activer.
La proposition d’une évolution
épigénétique par sélection de synapses a donné
lieu à de nombreux débats sur la possibilité qu’une
sorte de « darwinisme neural » poursuive en quelque
sorte l'évolution génétique des espèces biologiques,
mais au niveau de l'individu et, par conséquent, à celui du groupe
social.
Certains, comme le biologiste Richard Dawkins,
ont par exemple donné le nom de « mèmes
» à ces états cognitifs sélectionnés et transmissibles
d’un individu à l’autre, une forme de vie à part entière
dont le substrat serait les cerveaux humains.
Au cours du développement,
bien avant que les facteurs de croissance neuronaux soient sécrétés,
un grand nombre d'inductions moléculaires surviennent.
C’est le cas lorsqu’un un groupe de cellules influence le développement
d'un autre groupe cellulaire en sécrétant des substances dites morphogènes.
La formation de la
plaque neurale à partir de l’ectoderme est par exemple induite
par des cellules du mésoderme.
Ces facteurs inducteurs, qui possèdent
pour la plupart des similitudes avec les facteurs de croissance, agissent en stimulant
des récepteurs de la membrane cellulaire, ce qui influence l’activité
de certains gènes impliqués dans la différenciation cellulaire.
Différentes substances inductrices peuvent être sécrétées
successivement pour induire différentes étapes de la différenciation.
Ou encore, une même molécule inductrice peut être diffusée
dans le milieu et créer un gradient de concentration, à la manière
des facteurs de guidage chimiotrophiques.
L’exposition à différentes concentrations d’une même
substance peut aussi déboucher sur des différenciations cellulaires
distinctes.
FORMATION ET STABILISATION SÉLECTIVE DES
SYNPASES
Quand le cône de
croissance d’un axone entre en contact avec sa cible, il s’aplatit
et forme une synapse
avec elle. C’est à la jonction
neuromusculaire que le mécanisme de formation des synapses est le plus
connu.
On sait par exemple que des protéines sécrétées
par le cône de croissance lui-même contribuent à mettre en
place les récepteurs sur la membrane post-synaptique du muscle à
l’endroit précis où le cône de croissance vient en contact
avec lui. L’une de ces protéines est l’agrine
qui se fixe sur un récepteur particulier appelé MuSK (pour «
Muscle-specific kinase », en anglais) qui se retrouve un peu partout sur
la membrane cellulaire du muscle.
Les récepteurs MuSK sur lesquels
se fixe l’agrine vont rendre active une protéine intracellulaire
dénommée rapsyne qui elle va conduire à
l’agrégation des récepteurs
nicotiniques cholinergiques sur la membrane musculaire exactement sous le
cône de croissance. Des travaux plus récents montrent cependant que
le muscle n’aurait pas besoin du cône de croissance pour commencer
lui-même le travail d’agrégation des récepteurs, contribuant
ainsi à déterminer l’emplacement des plaques
motrices.
Parallèlement, des facteurs sécrétés
par la cellule cible, dans ce cas-ci le muscle, vont stimuler l’entrée
de calcium dans le cône de croissance. Celui-ci aura plusieurs effets sur
la terminaison présynaptique, dont celui de modifier le cytosquelette pour
donner sa forme aplatie au bouton présynaptique. L’entrée
de calcium va également favoriser la libération de neurotransmetteurs.
Cette transmission
synaptique rudimentaire va donner lieu, quelques semaines plus tard, à
une synapse capable de transmettre l’activité nerveuse.
Sur les muscles striés des vertébrés, les récepteurs
cholinergiques sont confinés dans la région de la plaque motrice.
La densité des récepteurs atteint à la plaque motrice 20
000 récepteurs par micromètre carré alors qu’ailleurs
sur la fibre musculaire elle est d’à peine 50. Or si l’on coupe
le nerf moteur et qu’on le laisse dégénérer, la distribution
des récepteurs devient plus dense un peu partout sur la fibre, sans toutefois
atteindre la densité initiale sous la plaque motrice. D’autre part,
si le muscle est innervé à nouveau, les récepteurs apparus
hors de la plaque motrice disparaissent. Seuls persistent ceux au niveau de la
plaque motrice.
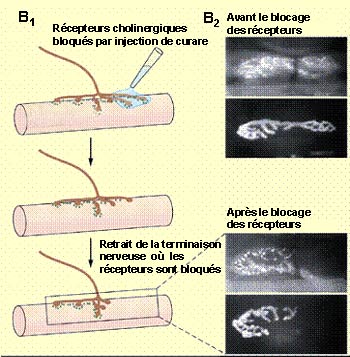
Des
expériences montrent également qu’en bloquant les récepteurs
de l’acétylcholine de la jonction neuromusculaire avec un antagoniste
comme le curare, l’innervation polyneuronale persiste. De même, si
l’émission de potentiels d’action des motoneurones est bloquée
avec un bloqueur de canaux sodiques comme la tétrodotoxine, la réduction
de l’innervation n’a pas lieu.
Ces expériences montrent
bien que le processus par lequel différentes fibres nerveuses rivalisent
entre elles pour s’approprier une cellule cible particulière est
régulé par l’activité électrique des partenaires
pré- et post-synaptiques.
Les
stades subséquents du développement de la synapse dépendront
justement de l’activité nerveuse. Celle-ci va influencer de façon
significative les interactions compétitives entre neurones, interactions
qui ne se feraient pas dans le sens d’un rejet actif de la terminaison nerveuse
perdante par la gagnante. Au contraire, on assisterait plutôt à une
perte progressive des structures post-synaptiques spécialisées associées
à certaines terminaisons nerveuses, perte qui diminuerait l’activité
électrique entre cette terminaison et le muscle, favorisant d’autant
plus la perte de récepteurs synaptiques. Ce processus ferait se rétracter
et s’atrophier naturellement les terminaison nerveuses ainsi désavantagées.
Parallèlement
à l’ajustement
de la taille des populations de neurones pré- et postsynaptiques, la
réorganisation synaptique représente donc l’étape finale
de la mise en place des circuits cérébraux. Et contrairement aux
étapes précédentes l’activité neuronale et la
transmission synaptique jouent donc un rôle primordial dans cette sculpture
fine des réseaux neuronaux.
Le fait que des mécanismes de
compétition et d’élimination de synapses surviennent au cours
du développement était connu depuis plusieurs décennies mais
l’idée que cette élimination pouvait avoir lieu de manière
épigenétique, sous le contrôle de l'activité
du réseau et sur le mode " darwinien " a été d’abord
articulée par Jean-Pierre Changeux et son équipe en 1972.
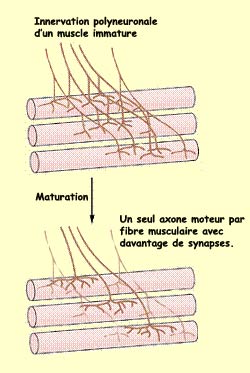
Les
travaux pionniers de Changeux et Danchin ont révélé qu’au
début du développement, les fibres musculaires sont innervées
par plusieurs axones mais ne conservent cependant à l’état
adulte que l’axone d’un seul motoneurone.
C’est aussi le cas des cellules
de Pukinje du cervelet qui ne le sont que par une seule fibre grimpante ou
encore des neurones
du cortex visuel qui reçoivent d’abord une innervation binoculaire
qui sera par la suite réduite en une ségrégation des afférences.
Les mammifères ont deux types
de fibres musculaires qui peuvent être identifiées par leur couleur
et leur vitesse de contraction : celles à contraction rapide sont pâles
et celle à contraction lente sont foncées. Les premières
sont impliquées dans des contractions phasiques alors que les autres le
sont davantage dans les ajustements posturaux.
On a aussi découvert
que les motoneurones qui innervent les muscles rapides conduisent l’influx
nerveux très rapidement et peuvent émettre des potentiels d’action
à des fréquences élevées de 30 à 60 influx
par seconde. À l’opposé, les motoneurones innervant les muscles
à contraction lente conduisent l’influx plus lentement et ne peuvent
générer que de 10 à 20 potentiels d’action par seconde.
Il y a donc une correspondance claire entre un type de motoneurone et un type
de fibre musculaire.
Or on sait que les chatons nouveau-nés n’ont
par exemple que des fibres musculaires à contraction lente et que celles-ci
vont se différencier en fibres lentes et rapides durant ses premières
semaines de vie. On peut alors se demander si ce sont les fibres musculaires qui
vont induire le bon type de motoneurone correspondant ou bien si ce sont plutôt
les neurones qui, en innervant les fibres musculaires, vont leur transmettre en
quelque sorte leur spécificité.
Des expériences
où l’on inversait chirurgicalement des motoneurones rapides sur des
fibres lentes et vice-versa ont permis de montrer que c’était cette
dernière hypothèse qui était la bonne: c’est le système
nerveux qui contrôle la rapidité de contraction de la fibre musculaire
qu’il va innerver. Cela implique donc que les motoneurones sont capables
d’influencer l’expression des gènes de la fibre musculaire
puisque la vitesse de contraction d’une fibre musculaire dépend de
versions différentes des chaînes légères des fibres
de myosine.
Ce phénomène remarquable pourrait s’établir en partie
à cause du pattern d’activité différent des deux types
de motoneurone qui aurait un effet de différenciation distinct sur les
fibres musculaires.