|
|
| Un livre accompagne le site depuis octobre 2024. Découvrez-le ici > | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|

|
|
Voir ce qu’il y a dans notre oeil Notre cerveau peut faire disparaître des objets immobiles Se mettre à jour avec « l’École des profs » et faire avancer les neurosciences avec Mozak Une nouvelle voie neuroanatomique au cortex visuel (plus deux outils pédagogiques)
|
|
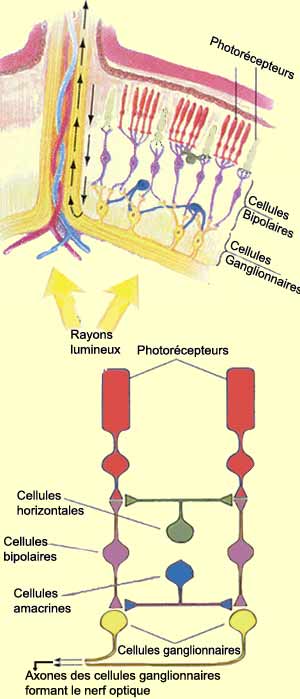
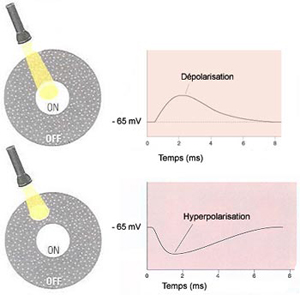
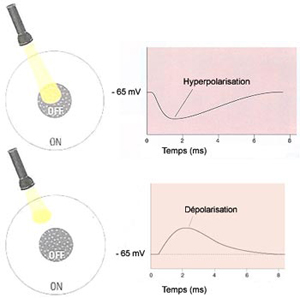
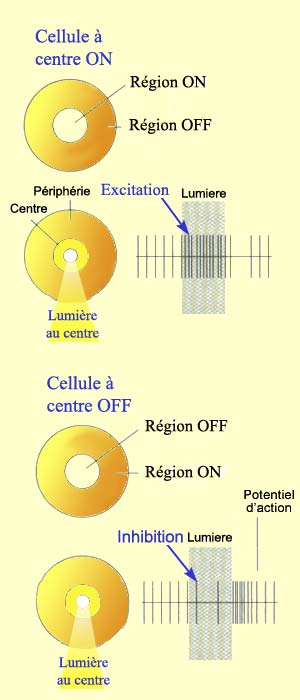
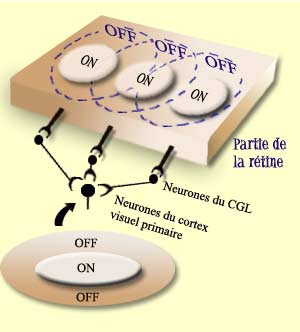
Pour voir, il faut d’abord que l’œil forme une image précise de la réalité sur la rétine. Il faut ensuite que l’intensité lumineuse soit transformée en influx nerveux par les cellules photoréceptrices de cette rétine.
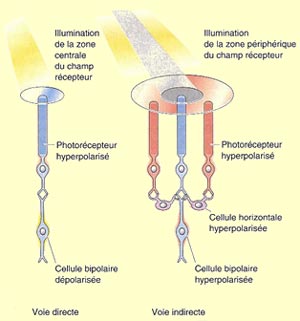
À côté de cette voie directe
qui va des photorécepteurs au cerveau, deux autres types
de cellules participent au traitement de l’information visuelle
dans la rétine. D’une part les cellules horizontales
reçoivent de l’information des photorécepteurs
et la transmettent à plusieurs neurones bipolaires environnants.
Et d’autre part les cellules amacrines reçoivent
leurs inputs des cellules bipolaires et procèdent de la même
façon avec les neurones ganglionnaires c’est-à-dire
activent ceux qui sont dans les environs. |
| |||||||||||||||||||||||||||||||||||||||||
|
|
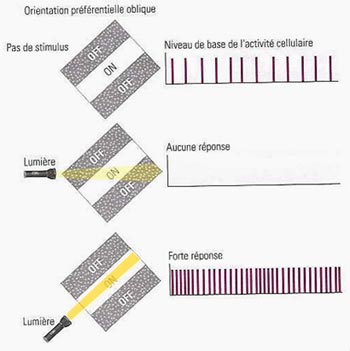
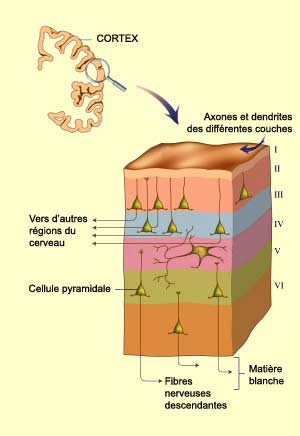
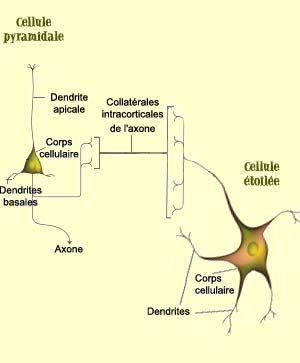
La couche IV contient par exemple de nombreuses cellules étoilées, de petits neurones avec des dendrites disposées en rayon autour du corps cellulaire. Les cellules étoilées reçoivent des connexions en provenance du corps genouillé latéral. Il s’agit donc d’une couche plutôt spécialisée dans la réception de l’information.
Pour sa part, la couche 1 contient très
peu de neurones. Elle est formée des axones et des dendrites des cellules
des autres couches. |
| |
|
|
|
|
|
|
|
|