Un livre accompagne le site depuis octobre 2024. Découvrez-le ici >
De
l'embryon à la morale
Les
100 000 milliards de cellules du corps humain adulte
sont réparties en 300 sortes de cellules différentes
où ne s’exprime qu’un sous-ensemble
du génome propre à un type particulier de
cellule.
Étonnamment, seulement 1,1 % de l’ADN de nos gènes
correspond aux
plans de fabrication de protéines. Comment alors
fabriquer les dizaines de milliers de protéines d’un
être humain si l’on ne possède qu’autour
de 30 000 gènes ? De toute évidence, le modèle
«un gène, une protéine» ne tient
plus.
La réponse réside dans les processus dits «post-transcriptionnels».
Par exemple, le «gène de l’insuline»,
cette hormone du métabolisme du sucre, code pour la
fabrication d’une très grosse protéine
qui sera par la suite découpée en plus petits
bouts par des enzymes. Et c’est la réunion de
deux d’entre eux (on dit aussi l’épissage)
qui donne l’insuline. En tout, ce seul gène sert
à produire quatre protéines et trois hormones.
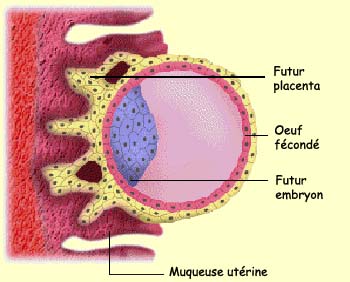
Quand l’embryon
devient profondément imbriqué dans l’utérus,
des ouvertures se forment dans la couche de cellules externes
et se remplissent de sang maternel, premier contact de l’embryon
avec la source de nutriment qui l’accompagnera tout au
long de son développement. Le
placenta est donc formé à la fois à
partir des trophoblastes embryonnaires et des cellules endométriales
de l’utérus de la mère.
Des remaniements hormonaux importants se mettent aussi en place
suite à l’implantation. L’hormone gonadotrophine
chorionique humaine (hCG) est sécrétée
par les trophoblastes de l’embryon et va stimuler la production
de progestérone et d’œstrogène par
les ovaires. C’est la hCG que les tests de grossesse détectent.
Du 10e au 14e jour, la cavité amniotique (qui
protégera le fœtus des coups) et le placenta (lieu
d’échange entre le fœtus et la mère)
commence à se former. À ce stade, la femme vient
tout juste de s’apercevoir qu’elle n’a pas
eu ses règles…
L’implantation se
fait normalement autour du 10e jour dans le tiers supérieur
de l’utérus, sur la ligne médiane. Lorsque
l’embryon s’implante ailleurs que dans l’utérus,
c’est le plus souvent dans les trompes de Fallope. On
parle alors de grossesses ectopiques pour
désigner ces implantations inadéquates.
DE LA FÉCONDATION À L'EMBRYON
La reproduction sexuée à l’œuvre chez
l’humain est rendue possible par la fusion de deux cellules
reproductrices appelées gamètes, l’ovule
maternel et le spermatozoïde paternel. Cette fusion va
permettre la formation d'un embryon contenant 46 chromosomes, dont
la moitié provient de la mère et l'autre moitié du
père. L’identité génétique singulière
qu’acquiert ainsi l’embryon fait qu’on parle
ici bien davantage d'une procréation (formation d'un nouvel être)
que d'une reproduction
à l'identique.
Une journée après la
fécondation, le zygote entreprend une série
de divisions
mitotiques par lesquelles le volume considérable de
cytoplasme de l’ovule est réparti en plusieurs cellules
filles. Il n’y a donc pas de croissance de taille durant
ces premières divisions cellulaires qui font passer l’embryon
successivement au stade de 2, 4, 8, puis 16 cellules qu’on
appelle blastomères. On parle de segmentation pour
désigner cette première phase du développement
embryonnaire caractérisée par une suite de divisions
rapides et rapprochée.
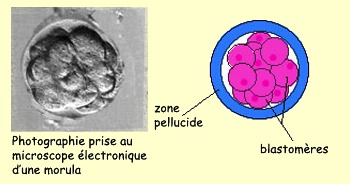
Le stade de 16 cellules, aussi appelé morula,
est atteint au troisième jour après fécondation
chez l’humain. À ce stade, l'embryon est une boule
compacte de cellules qui descend dans les trompes de Fallope vers
l'utérus. Les divisions n'ont toujours pas entraîné d'augmentation
de la taille de l'embryon par rapport à l'ovule initial.
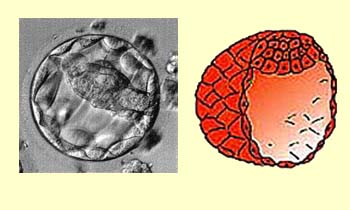
Au stade de blastula, à partir de 128
cellules, la sphère de cellules se creuse d'une cavité appelée
blastocoele. Ce stade survient vers le quatrième jour et
jusqu'à la fixation de l'embryon sur la paroi de l'utérus,
vers le sixième ou septième jour. C’est pourquoi
l’enveloppe protectrice (ou zone pellucide)
autour de la sphère de cellules va graduellement dégénérer,
ce qui va permettre la fixation de l'embryon aux parois de l'utérus.
C’est également à ce stade que se différencient
des cellules externes ou trophoblastes, qui contribueront
au placenta, et des cellules internes ou embryoblastes,
qui formeront l’embryon en tant que tel.
L’implantation (ou nidation)
dans la paroi utérine commence vers le septième jour
quand les cellules externes de l’embryon (ou trophoblastes),
débarrassées de la zone pellucide, sécrètent
une enzyme qui permet à l’embryon de s’enfoncer
dans la paroi de l’utérus. À mesure que les
trophoblastes prolifèrent, ils forment deux couches de cellules
distinctes : une qui demeure autour de l’embryon, et l’autre
dont les membranes cellulaires fusionnent ensemble en une masse
multinucléée appelée syncytium et
qui est responsable de l’invasion dans la paroi utérine.
La troisième semaine de développement débute
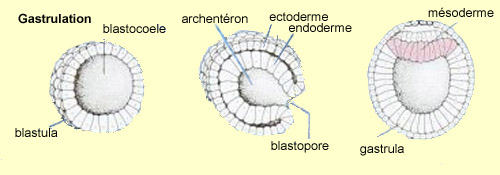
par une réorganisation cellulaire importante : la gastrulation.
La blastula va subir une invagination, c'est-à-dire qu'une
portion des cellules de surface de la sphère va pénétrer à l'intérieur
de celle-ci, formant ainsi l’endoderme, tandis que les cellules
demeurant
à l’extérieur vont former l’ectoderme.
Les cellules à l'intérieur de la sphère vont
ensuite se diviser en deux feuillets de cellules formant deux disques
superposés. Celui du dessus deviendra l’embryon alors
que celui du dessous se transformera en un sac vitellin fournissant
des éléments nutritifs en attendant un système
circulatoire fonctionnel.
Au début de cette phase, une étroite rangée
de cellule trace un sillon sur le disque embryonnaire et définit
ainsi l’axe général autour duquel se développera
notre corps bilatéral. On peut officiellement parler du
début de l’embryon en tant que tel à
partir de ce stade où une troisième couche de cellules,
le mésoderme, vient se glisser entre les
deux déjà présentes, l’ectoderme et
l’endoderme.
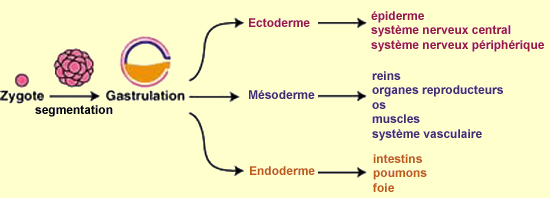
Toutes les cellules de ces couches ont le même matériel
génétique, mais certaines commencent à
exprimer certains gènes plutôt que d’autres
afin de développer différents organes. L’endoderme,
la couche la plus intérieure, produira entre autres les
intestins, les poumons et le foie. Le mésoderme,
la couche mitoyenne, donnera naissance aux reins, aux organes reproducteurs,
aux os, aux muscles et au système vasculaire. Et l’ectoderme,
la couche extérieure, sera à l’origine à la
fois de l’épiderme et de tout le système nerveux
central et périphérique.
Source : modifié à
partir de BrainConnection.com
Les mouvements coordonnés permettant la gastrulation, appelés mouvements
morphogénétiques, impliquent l'embryon entier.
Les cellules adoptent de nouvelles positions et ont par conséquent
de nouvelles voisines. La gastrulation permet donc de rassembler
des sous-groupes de cellules qui peuvent ainsi agir les unes sur
les autres par des
phénomènes d’induction.
En résumé, on peut dire que la gastrulation est
la période des 3 : 3ème semaine,
3 couches cellulaires à l’origine de 3 structures
importantes : la ligne primitive qui définit
le plan de symétrie bilatérale du futur embryon,
la corde dorsale (ou notochorde)
qui induira la formation de la plaque neurale lors
du stade suivant, la neurulation.
À partir de six
à huit semaines après la fécondation,
les hémisphères cérébraux commencent
à se développer. Autour de la septième
semaine, des nerfs font des connexions avec des muscles qui
permettent à l’embryon de faire des mouvements
spontanés.
À la fin de la huitième semaine, toutes les structures
essentielles internes et externes sont présentes. Le
second et troisième trimestre seront essentiellement
consacrés à la croissance de ce qui vient d’être
mis en place.
Dans l’épithélium
du tube neural, la
prolifération a lieu à des endroits spécifiques
appelés zones germinales. Pour la plupart des régions
du système nerveux, ces zones germinales sont situées
près de la surface de l’une des cavités
qui deviendront le
système ventriculaire du cerveau.
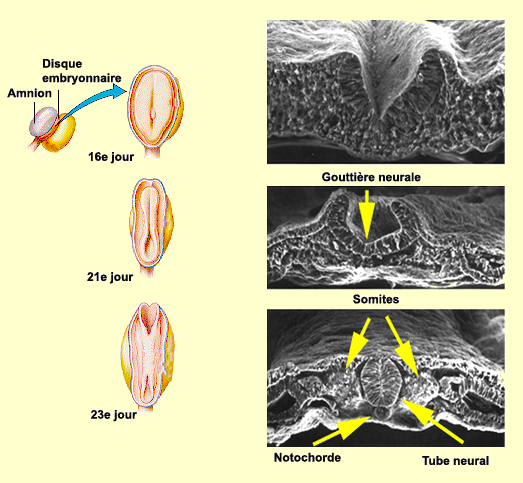
LA MISE EN PLACE
DU SYSTÈME NERVEUX
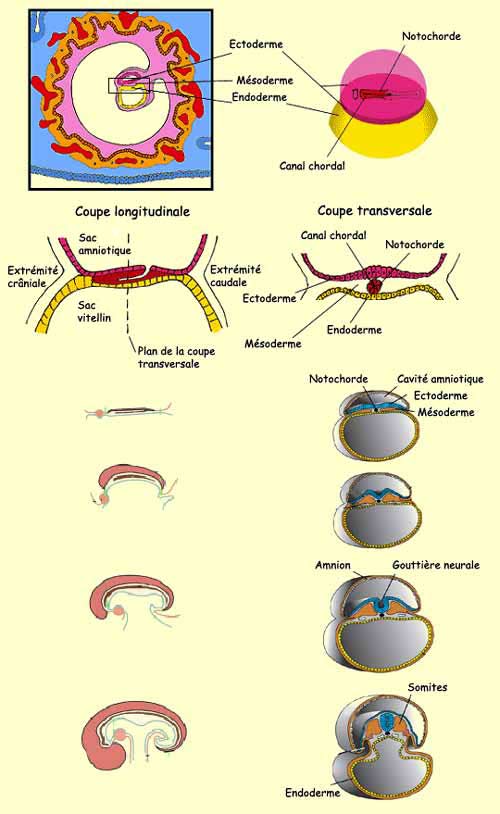
Environ 3 semaines après sa conception, le cerveau humain
n’est qu’une simple couche de cellules aplaties de
l’ectoderme appelée plaque
neurale. On assiste par la suite à la formation
d’un sillon qui s’étend de la partie rostrale à la
partie caudale de cette plaque. Les parois de ce sillon
neural vont ensuite former une gouttière
neurale dont la fermeture, d’abord en son milieu
puis dans sa partie antérieure et postérieure,
va former le tube neural. Des cellules de la
partie dorsale de ce tube deviendront quant à elles la crête
neurale, structure à l’origine de neurones du
système nerveux périphérique.
La partie de la plaque neurale située juste au-dessus
de la corde se différencie en plaque du plancher. Les
signaux inducteurs en provenance de cette plaque du plancher induisent
le développement des motoneurones
spinaux et bulbo-pontiques à partir des cellules les
plus ventrales du tube neural. Les cellules les plus éloignées
dorsalement donneront pour leur part les neurones sensitifs.
Le processus de formation du tube
neural, qui débute
souvent avant même que la mère sache qu’elle
est enceinte, est nommé neurulation.
C’est à partir de ce tube que se développeront
le cerveau et la moelle épinière qui sont alors
les organes les plus développés de l’embryon
et qui lui donne sa forme incurvée caractéristique.
À la fin de la troisième semaine, les yeux et
les oreilles auront aussi commencé à se former.
Après la
segmentation et la gastrulation,
on rentre donc dans la phase de l’organogenèse qui
va aboutir à la mise en place des groupes de cellules
qui deviendront les différents organes du corps humain.
On parle de métamérisation pour
décrire les divisions du mésoderme en une suite
de segments identiques, les métamères, qui se
répètent le long de l'axe longitudinal de l’embryon.
Le développement du mésoderme forme à ce
stade des bourgeonnements appelés somites de
chaque côté du tube neural. C’est à partir
de ces somites que vont se développer les 33 vertèbres
de notre colonne vertébrale et les muscles squelettiques
correspondants.
Source : Dr. K. Tosney, University
of Michigan
Au début de la 4ème semaine après la fécondation,
le tube neural se ferme complètement, achevant la première étape
de développement du cerveau et de la moelle épinière.
L’étape suivante, l'histogenèse,
c’est-à-dire la différenciation
cellulaire à partir de cellules souches qui mène
à la formation des tissus nerveux, va pouvoir commencer
pour de bon. Elle se fera en parallèle avec la
formation des grandes subdivisions du cerveau et le réarrangement
des populations cellulaires qui s’ensuit.
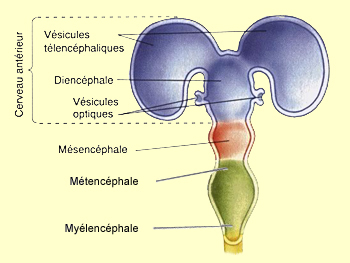
Le renflement le plus
rostral, le télencéphale, voit deux bourgeonnements
jaillir de sa partie antérieure. Ces deux vésicules
télencéphaliques prendront rapidement de l’ampleur
pour former les deux hémisphères cérébraux,
en s’agrandissant d’abord postérieurement
au-dessus du diencéphale, puis sur ses côtés.
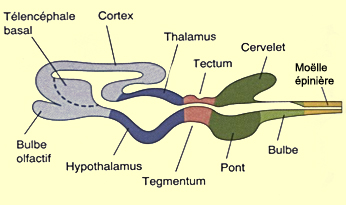
Schéma général
du cerveau en coupe sagittal (valable pour tous les mammifères)
Une autre paire de vésicules
va également bourgeonner sur la surface ventrale de
ces hémisphères cérébraux pour
donner naissance aux bulbes olfactifs et aux
autres structures qui contribuent à l’olfaction.
Diverses structures émergeront par la suite à
partir des parois du télencéphale pendant que
la substance blanche reliant ces structures se développera
aussi. Les neurones de la paroi du télencéphale
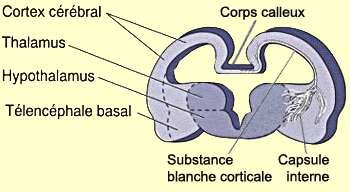
prolifèrent et forment donc trois régions distinctes
: le cortex cérébral, le télencéphale
basal et le bulbe olfactif.
Les axones de ces neurones
vont aussi s’allonger progressivement pour communiquer
avec les autres parties du système nerveux. Certains
constitueront la substance blanche corticale qui
part de neurones du cortex ou s’y projette. D’autres
formeront le corps calleux, ce pont axonal
entre les deux hémisphères. D’autres
enfin, ceux de la capsule interne, relieront
la substance blanche corticale au tronc cérébral
généralement via le thalamus. Les
axones des neurones moteurs du cortex passeront par exemple à travers
la capsule interne pour rejoindre les motoneurones de la
moelle épinière.
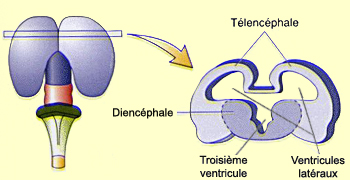
L'espace restant entre le télencéphale et le
diencéphale donne naissance aux ventricules
cérébraux. L’espace situé
au centre du diencéphale forme le troisième ventricule.
Les deux ventricules latéraux sont aussi appelés
le premier et deuxième ventricule.
Le diencéphale se différencie également
en deux territoires distincts : le thalamus et
l’hypothalamus.
De chaque côté
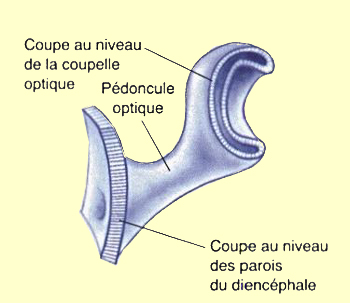
du diencéphale se développent aussi deux vésicules
secondaires, les vésicules optiques.
Les vésicules optiques s’allongent et se replient
vers l’intérieur pour former les pédoncules
et les coupelles optiques qui donneront naissance à la
rétine et au nerf
optique. La rétine et le nerf optique ne font donc
pas partie du système nerveux périphérique,
mais appartiennent bel et bien au cerveau !
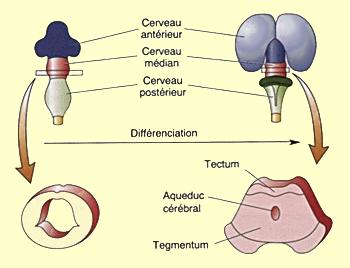
La vésicule mésencéphalique se transforme
beaucoup moins que la vésicule prosencéphalique.
Sa surface dorsale forme le tectum et
son plancher le tegmentum. Durant la différenciation
de ces structures, l’espace qui les sépare
se rétrécit en un
étroit canal qui forme l’aqueduc cérébral.
Dans sa partie rostrale, celui-ci débouche dans
le troisième ventricule du diencéphale.
C’est à travers le mésencéphale que
passeront les faisceaux de fibres qui relient le cortex et la moelle,
tant ceux qui montent en provenance du système sensoriel
que ceux qui descendent pour contribuer au contrôle
du mouvement.
Le tectum se différencie en deux structures qui reçoivent le nom
de colliculus. Le colliculus
supérieur reçoit directement des informations
de l'oeil et contrôle les mouvements oculaires alors que le colliculus
inférieur reçoit les informations provenant des oreilles
et constitue un relais important dans les voies auditives.
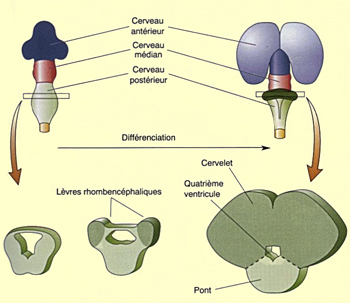
La partie
rostrale du cerveau postérieur, le métencéphale,
se différencie en deux structures importantes, le cervelet et
le pont. Le cervelet naît
de l’épaississement du tissu qui tapisse les
parois latérales du tube neural à cet endroit.
Les deux masses ainsi formées finissent par fusionner
dorsalement pour former le cervelet. Pendant ce temps, un
renflement se forme sur la partie ventrale du métencéphale
pour former le pont (aussi appelée le protubérance).
Cette structure est une voie de passage importante de l’information
entre cerveau, cervelet et moelle
épinière.
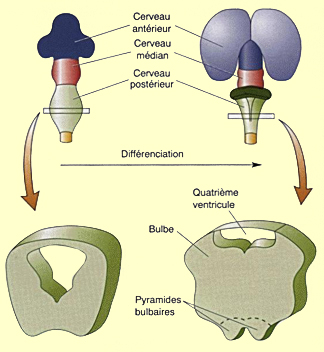
Des changements moins spectaculaires surviennent dans
la partie caudale du cerveau postérieur, le myélencéphale,
avec le renflement des régions ventrales et latérales
de cette structure qui donnera naissance au bulbe
rachidien. Le long de la surface ventrale du bulbe
se développeront aussi les deux
pyramides bulbaires formées par le passage des
faisceaux corticospinaux responsables de la motricité
volontaire. Enfin le canal central, qui persiste au cours
de la formation du bulbe, devient le quatrième ventricule.
Tout le reste du tube neural
situé caudalement par rapport aux cinq grandes vésicules
va se transformer en moelle
épinière. Cette différenciation
se fera de manière assez directe par l’épaississement
des parois du tube. Cela va progressivement diminuer le diamètre
du tube neural qui deviendra le minuscule canal spinal.
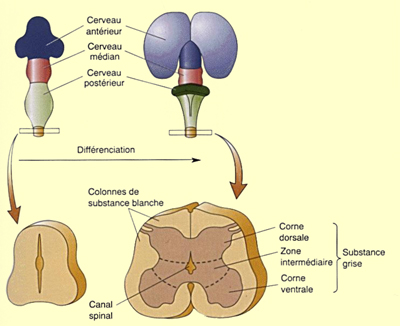
En coupe transversale, on observe que les corps neuronaux se
regroupent dans la
partie centrale grise de la moelle (en forme de
papillon) tandis que la zone périphérique
blanche est formée de faisceaux d'axones.
La matière grise de la moelle épinière peut être à son
tour divisée en corne dorsale qui reçoit
les afférences sensorielles et en corne ventrale dont
les neurones vont innerver les muscles squelettiques. De même,
dans la substance blanche, se développent des colonnes
dorsales formées d'axones sensoriels remontant
vers le cerveau et des colonnes latérales faites
d’axones du faisceau cortico-spinal qui descendent pour transmettre
des signaux de contrôle du mouvement. Entre la corne dorsale
et ventrale vont aussi se développer un grand nombre de
neurones intermédiaires impliqués dans différents
types de réflexes et l'établissement de réseaux
permettant un premier traitement de l’information reçue
au niveau de la moelle épinière.