La mitose est
le type de division cellulaire qui s’opère dans
les cellules de notre corps que ce soit durant le développement
de celui-ci ou durant le renouvellement des tissus chez l’adulte.
La cellule qui entre en mitose passe par plusieurs étapes
qui aboutissent à sa division en deux cellules filles
semblables à la cellule mère. Les cellules
filles peuvent ensuite se diviser à leur tour, ce
qui aboutit
à une croissance exponentielle du nombre de cellules
durant le développement embryonnaire.
La méïose est l’autre grand
type de division cellulaire réservé exclusivement
aux gamètes (ovules et spermatozoïdes). Spécifique
aux eucaryotes,
la méïose survient lorsqu’une cellule mère
diploïde produit quatre cellules filles haploïdes
ayant toutes un génome différent.
Éventuellement, une de ces cellules haploïdes mâle
va fusionner avec une cellule haploïde femelle pour redonner
une cellule diploïde lors de la
fécondation.
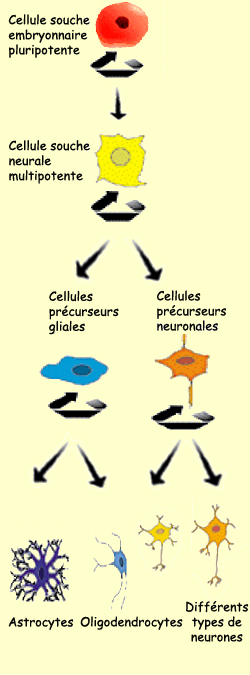
Les cellules souches,
en se divisant, peuvent produire soit deux autres cellules
souches, soit deux neuroblastes, soit une cellule souche
et un neuroblaste. Dans le premier cas, les cellules souches
filles conservent la propriété de produire
d’autres cellules souches pluripotentes.
On parle de différenciation lorsqu’une cellule
souche donne naissance à au moins un neuroblaste parmi
ses deux cellules filles. Une cellule différenciée
possède une forme et une fonction plus spécialisée
que la cellule mère. Elle ne peut donc produire que
les cellules de sa lignée particulière et ne
peut pas revenir en arrière pour donner naissance à
des cellules souches.
Le processus général
qui conduit à la naissance de nouveaux neurones est
appelé neurogenèse. La plupart
des neurones du néocortex sont formés entre
la cinquième semaine et le cinquième mois de
gestation chez l’humain.
Même si l’essentiel du développement neuronal
du cortex intervient avant la naissance, certaines parties
du cerveau humain adulte conservent la capacité de produire
de nouveaux neurones (voir capsule histoire ci-bas).
Cette neurogenèse dans le cerveau adulte représente
un processus trop limité pour remplacer des populations
de neurones qui auraient été détruites
par des lésions ou des maladies. On espère néanmoins
que la compréhension des processus de régulation
de cette neurogenèse, en particulier le
rôle des facteurs environnementaux comme le stress,
nous donne des pistes pour préserver la croissance de
nouveaux neurones en réponse à une dégénérescence
pathologique.
DES CELLULES SOUCHES AUX NEURONES
En quelques mois seulement, la totalité de
nos 100 milliards de neurones (à
quelques exceptions près) et un nombre encore
supérieur de cellules
gliales seront produits à partir d’une petite
population de cellules précurseurs.
Les cellules souches qui prolifèrent dans la zone
ventriculaire du tube neural sont à l’origine
des deux grandes familles de cellules que sont les neurones
et les cellules gliales. Mais la différenciation cellulaire
ne s’arrête pas là. Les
différentes structures cérébrales sont
formées d’innombrables
types de cellules nerveuses qui se distinguent par leurs neurotransmetteurs,
les molécules à la surface de leur membrane,
le type de synapses qu’elles forment et reçoivent,
etc.
Les
processus de prolifération, de détermination
et de différenciation cellulaire doivent donc
générer toute cette diversité
en procédant par étapes. À chacune de
celles-ci, le destin de la cellule se précise. Concrètement,
cette maturation correspond à
des changements importants au niveau de la réplication
et de l’expression des gènes dans le noyau de
ces cellules.
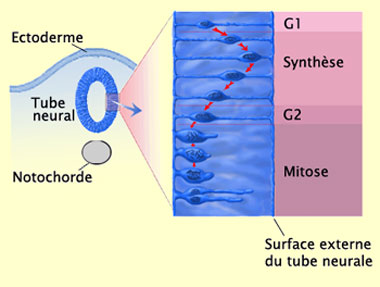
Lors de la phase de prolifération par exemple, les cellules
qui se divisent suivent les étapes du cycle classique des
divisions mitotiques (voir encadré) mais en les accompagnant
d’un mouvement oscillatoire assez particulier de leur noyau.
Celui-ci effectue un aller retour entre la zone dite ventriculaire
du tube neural (qui fait face au ventricule) et une autre dite
marginale (qui donne sur l’extérieur).
Les cellules nouvellement divisées
commencent par étendre un prolongement cytoplasmique jusqu’à
la surface externe du tube neural, du côté de la pie-mère
(G1, ou phase de croissance). Le noyau et le cytoplasme qui l’entoure
se déplacent à l’intérieur de cet étroit
cylindre de cytoplasme vers la surface externe.
Quand le noyau arrive à proximité de la surface externe,
il commence à répliquer son ADN (S, ou phase de syntèse
d'ADN). Le noyau migre ensuite à nouveau vers la lumière
du tube neural pendant que la cellule continue de se développer
(G2, ou phase de préparation à la mitose). La cellule
rétracte alors son prolongement à la surface externe
et entre en mitose (phase M).
Inspiré de : Crump Institute
for Biological Imaging
La
division cellulaire donne naissance soit à de nouvelles
cellules souches, soit à des cellules appelées
neuroblastes qui se différencieront en neurones (voir
l’encadré). Les premières reprendront le
cycle de la division en renvoyant à leur tour un prolongement
vers la surface externe du tube neural. Quant aux secondes,
elles quittent la zone ventriculaire et migrent
jusqu’à leur emplacement définitif dans
le cerveau en développement.
Une explication possible pour expliquer
cet étrange ballet serait la nécessité, pour
le noyau, d’être exposé selon une séquence
temporelle adéquate à différents facteurs cytoplasmiques
situés dans différentes régions à l’intérieur
de la cellule. Cette phase intense de prolifération cellulaire
donne lieu à une surproduction de neurones dont le nombre
sera ensuite réduit par apoptose (voir capsule à gauche).
Après leur dernière mitose dans la zone ventriculaire,
la plupart des cellules nouvellement formées entreprennent
une migration qui les mènera
à leur position définitive. Les
neuroblastes se guident de différentes façons
lors de leur migration. Dans certaines régions
comme le cortex ou le cervelet, la migration d’une
grande partie de ces neurones est facilitée par des
cellules gliales radiales qui étendent leur prolongement
de la zone ventriculaire jusqu’à
la surface corticale.
La structure, la fonction
et l’activité
d’une cellule dépendent en grande partie de ses
gènes. La séquence d’événements
cellulaires qui mène à la différenciation
des neurones est donc en partie contrôlée par
des facteurs intrinsèques. En d’autres
termes, des mécanismes cellulaires qui activent et désactivent
des gènes. L’acide rétinoïque, un
dérivé
de la vitamine A active par exemple des récepteurs spécifiques
qui modulent l’expression de certains gènes.
Mais chez les eucaryotes,
l’expression des gènes (ou phénotype) est
toujours influencée en plus par des facteurs
extrinsèques (on dit aussi épigénétiques)
venant de l’environnement de la cellule.
Des expériences de transplantation
ont ainsi montré que si les cellules greffées
sont prises sur un animal dont le développement est
assez avancé, les cellules se développeront
sur le nouvel hôte en gardant leur phénotype
original. Mais plus l’animal où est prélevé
le tissu est jeune, plus les cellules se développeront
sur le nouvel hôte en en adoptant le phénotype
de celui-ci, indiquant ainsi l’importance de l’influence
des facteurs extracellulaires de son nouvel environnement.
Pour comprendre le développement
du système nerveux, il faut donc constamment examiner
comment les signaux intrinsèques et extrinsèques
se combinent pour mener à
bien la détermination et la différenciation.
Certaines cellules nerveuses
comme les motoneurones attendent
d’être arrivés à leur emplacement
final avant d’étendre leurs prolongements axonal
et dendritiques. D’autres, comme les
cellules granulaires du cervelet, développent
leurs prolongements en même temps que leur migration
(voir l’encadré sous le texte).
CONNECTER LES NEURONES ENTRE EUX
L’endroit où
est situé chaque neurone dans notre cerveau est plus
critique que celui des autres cellules de notre corps. En
effet, les fonctions nerveuses dépendent des connexions
précises entre les neurones et leurs cibles. En d’autres
termes, les éléments pré et post-synaptiques
doivent être à la bonne place au bon moment.
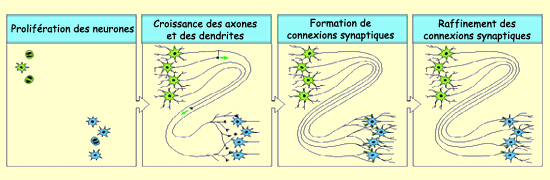
Après la phase de prolifération,
les cellules précurseurs des neurones quittent la lumière
du tube
neural et migrent vers
leur emplacement définitif dans le cerveau. Une fois
leur destination finale atteinte, le corps cellulaire du neurone développe
ses dendrites et son axone qui vont lui permettre de faire des connexions
avec les autres neurones.
Pour ce faire, les axones doivent trouver leur cible spécifique, ce qui
n’est pas une mince tâche. On peut considérer que la formation
de ces voies axonales se fait en trois temps : d’abord sélectionner
le bon trajet, ensuite choisir la bonne cible, et finalement s’établir
à la bonne destination.
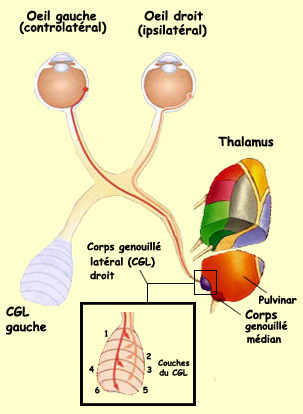
Prenons par
exemple l’axone d’une cellule
ganglionnaire de la rétine qui doit atteindre
le corps
genouillé latéral du thalamus. Cet axone
suit d’abord le nerf optique mais très vite
il atteint le chiasma
optique où trois possibilités s’offre
à lui : prendre le tractus optique droit, le tractus
optique gauche, ou encore suivre le nerf optique controlatéral.
Selon que la cellule ganglionnaire vient de la partie nasale
ou temporale de la rétine, elle devra sélectionner
le bon trajet c’est-à-dire le tractus
optique controlatéral ou le tractus optique ipsilatéral.
Après avoir atteint le thalamus, l’axone n’est
pas au bout de ses peines puisqu’il a le choix entre
une bonne douzaine de cibles possibles. Il devra donc choisir
la bonne cible ce qui veut dire dans ce cas-ci le
corps génouillé
latéral (CGL) et non le corps genouillé
médian du thalamus ou le pulvinar.
L’axone devra enfin s’établir
à la bonne destination finale de son parcours,
en l’occurrence ici la bonne couche du corps genouillé
latéral et la position rétinotopique adéquate.
Une fois qu’il a atteint sa cible,
l’axone va
élaborer une multitude de synapses avec elle. Puis un
processus de sélection dépendant de l’activité
des neurones va diminuer le nombre de ces synapses pour
ne conserver que celles qui jouent un rôle significatif
dans les circuits nerveux. Source : Dr. Brian E. Staveley, Department
of Biology, Memorial University of Newfoundland
Longtemps après la naissance, ce mécanisme de
renforcement synaptique lié à l’activité
neuronale continue à influencer nos synapses pour adapter
le corps à l’activité et aux perceptions
du monde extérieur.
Certains des mécanismes utilisés dans les premiers
stades du développement embryonnaire vont même s’adapter
pour participer aux modifications que le cerveau connaît
en permanence. Cette plasticité,
qui nous permet de nous adapter aux conditions changeantes de
notre environnement, est considérée comme
à la
base de notre mémoire.
Les premiers neurones à
être formés dans la zone ventriculaire du cortex
cérébelleux sont les
cellules de Purkinje et de Golgi qui migrent aussitôt
vers la zone marginale. La zone ventriculaire donne aussi
naissance à des neurones précurseurs qui vont
migrer au-delà de la couche de cellules de Purkinje
pour former une seconde zone germinale appelée la
couche granulaire externe. C’est à partir de
cette couche que vont se développer les trois types
principaux d’interneurones du cervelet : les
cellules en corbeille, les cellules étoilées
et les cellules granulaires. La naissance de ces
interneurones coïncide avec l’allongement des
dendrites des cellules de Purkinje.
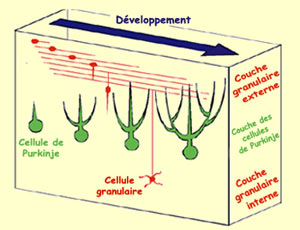
Pour atteindre leur destination
finale, les cellules granulaires doivent migrer d’abord à travers
la couche moléculaire, puis à travers
la couche de cellules de Purkinje. En arrivant dans
la couche moléculaire, les cellules granulaires
développent deux prolongements parallèles à la
surface du cortex cérébelleux et perpendiculaires
aux dendrites des cellules de Purkinje qui sont en
train de se développer. Un troisième
prolongement se forme bientôt et descend vers
la couche granulaire. Le corps cellulaire de la cellule
granulaire n’a plus qu’à suivre
ce prolongement pour atteindre sa position finale dans
la couche granulaire, laissant derrière lui
les deux prolongements qui forment les fibres parallèles
faisant des connexions aux ramifications dendritiques
des cellules de Purkinje.
Différenciation
des cellules granulaires et
des cellules de Purkinje dans le cervelet.
Source: Mineko KENGAKU, Laboratory for
Neural Cell Polarity