La destruction du cortex moteur
(par un accident cérébro-vasculaire par exemple) amène la
perte des mouvements précis, en particulier ceux des mains et des doigts.
L’apprentissage de nouveaux mouvements est peu affecté par des dommages
au cortex cérébral. La mémoire de séquences motrices
déjà apprises est aussi largement épargnée, bien que
l’exécution des mouvements est plus maladroite. Ces observations
montrent que c’est plutôt le cervelet
que le cortex qui joue un rôle important dans l’apprentissage et la
mémoire du mouvement ou mémoire
procédurale.
L’aire 4 du gyrus précentral
n’est pas la seule aire corticale contribuant au système pyramidal.
C’est cependant la zone corticale où l’on réussit à
produire des mouvements avec les stimulations électrique de plus basse
intensité. Autrement dit, des stimulations électriques qui ne sont
pas efficaces pour produire des mouvements à partir des autres aires corticales
le sont suffisamment pour déclencher des mouvements à partir de
l’aire 4.
LE CORTEX MOTEUR
Les structures cérébrales
impliquées dans la motricité sont nombreuses. Certains vont même
jusqu’à dire que c’est pratiquement tout le cerveau qui participe
au mouvement. Car même si le cortex moteur est habituellement associé
aux aires
4 et 6, le contrôle du mouvement volontaire implique en réalité
presque toutes les aires du néocortex.
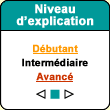
Le
cortex moteur primaire correspond pour sa part à la région anatomique
désigné par l’aire 4 du gyrus précentral. L'introduction
des opérations au cerveau vers le milieu du XXe siècle a permis
à certains neurochirurgiens comme le Dr. Wilder Penfield à Montréal
de confirmer l’emplacement du cortex moteur primaire. Au cours d’interventions
chirurgicales destinées à soulager des patients de leurs crises
d’épilepsie, Penfield stimule les régions corticales afin
de préserver les zones vitales de l’ablation. Il découvre
alors que des stimulations du gyrus précentral déclenche des contractions
musculaires très localisées du côté
controlatéral du corps et qu’il existe une représentation
somatotopique (voir encadré sous ce texte) des régions du corps
correspondantes sur le cortex moteur primaire (aire 4).
Penfield
montre également que l’aire corticale 6, située juste rostralement
par rapport à l’aire 4, possède quant à elle deux autres
représentations somatotopiques induisant des mouvements complexes lorsqu’on
les stimule. La première est dans la région latérale de l’aire
6 et se nomme aire prémotrice (ou APM). Elle contribue à
guider les mouvements en intégrant les informations sensorielles et s’occupe
des muscles qui sont les plus proches de l’axe du corps.
La seconde
représentation somatotopique se trouve dans l’aire motrice supplémentaire
(ou AMS) qui est située dans la partie médiane de l’aire
6. L’AMS est impliquée dans la planification de mouvements complexes
et dans la coordination de mouvements impliquant les deux mains.
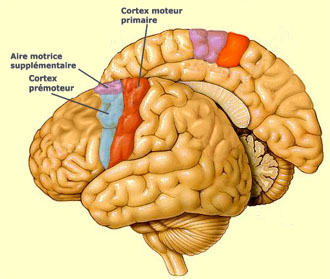
Les expériences de stimulation
corticale de Penfield ont permis de dresser une cartographie complète du
cortex moteur appelée "homoncule moteur" (car il y a d'autres
homoncules, comme le sensoriel par exemple). Le détail le plus frappant
de cette cartographie est que les surfaces allouées sur le cortex ne sont
pas proportionnelles à la taille de la partie du corps correspondante,
mais plutôt à la complexité des mouvements que cette partie
peut effectuer. Pour cette raison, les surfaces allouées à la main
et au visage ont une taille disproportionnée par rapport au reste du corps.
Pas étonnant puisque c'est justement la dextérité et la rapidité
de mouvement des mains et de la bouche qui confèrent à l'homme deux
de ses facultés les plus spécifiques: se servir d'outils et parler.
Les fonctions des ganglions de la
base sont complexes et encore peu connues. Les gens qui souffrent de la maladie
de Parkinson, caractérisée par des tremblements et une difficulté
à initier les mouvements, montrent un déficit de dopamine
dans les ganglions de la base. Comme ceux-ci contribuent d’une manière
importante à déterminer différents paramètres du mouvement,
leur mauvais fonctionnement entraîne les problèmes moteurs liés
à la maladie de Parkinson.
Des anormalités dans les ganglions
de la base se manifestent aussi chez les patients souffrant de la maladie d’Huntington
ou du Syndrome de la Tourette. Ceux-ci sont aux prises avec des mouvements involontaires
qui provoquent grimaces, spasmes et tics de toutes sortes.
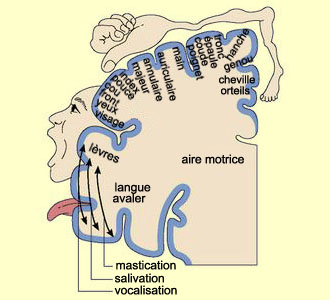
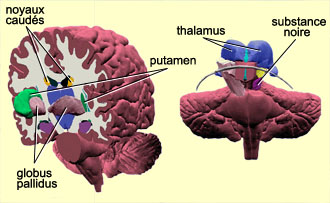
LES GANGLIONS DE LA BASE
Plusieurs structures cérébrales
sont regroupées sous l’appellation de ganglions de la base, soit
le noyau caudé, le putamen, le globus pallidus et le noyau sous-thalamique.
La substance noire, une structure mésencéphalique richement interconnectée
aux ganglions de la base, bien que ne faisant pas à proprement parler partie
de ces derniers, leur est souvent associée.
Les ganglions de la base sont impliqués
dans une boucle
complexe qui les lie à différentes aires corticales. L’information
en provenance des aires frontales, préfrontales et pariétales du
cortex traverse les ganglions de la base et retournent à l’aire motrice
supplémentaire via le thalamus. Les ganglions de la base exerceraient ainsi
une action facilitatrice sur le mouvement en focalisant les informations en provenance
de différentes régions corticales sur l’AMS. Ils sont aussi
susceptibles d’agir comme un filtre bloquant la réalisation des mouvements
lorsque ceux-ci sont inadaptés.
Plusieurs des circuits impliquant les ganglions de la base
ne sont cependant pas moteurs, mais sont plutôt impliqués dans la
mémorisation et le traitement des processus cognitifs et émotifs.
Les ganglions de la base sont encore largement méconnus et semblent jouer
un rôle beaucoup plus large que sa contribution au contrôle moteur.
Le cervelet est aussi une
machine à apprendre et à mémoriser grâce
à ses connexions nerveuses modifiables où tout ce qui y est programmé
est constamment comparé avec les résultats obtenus. Quand cette
comparaison ne permet pas de réaliser de façon satisfaisante ce
qui est attendu, l’activité cérébelleuse modifie de
façon compensatoire la séquence de mouvement pour rendre l’action
plus efficace. Cette mémoire
procédurale se constitue donc automatiquement avec la pratique,
sans l’aide d’un contrôle conscient.
Le cervelet jouerait également
un rôle majeur dans l’apprentissage de la coordination entre les différents
segments corporels. En effet, le déplacement de chacun des segments de
notre corps a un effet sur le suivant à cause de sa masse. Le cervelet
apprendrait donc à calibrer les commandes musculaires en terme de temps
et de force afin de corriger à l’avance l’effet de ces interactions
sur la trajectoire.
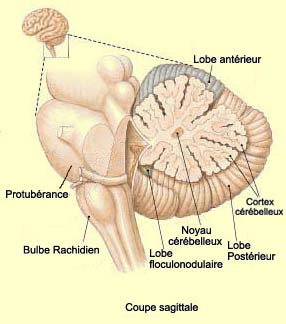
LE CERVELET
Le
cervelet semble jouer plusieurs rôles : il emmagasine des séquences
de mouvement apprises, participe à l’ajustement fin (« fine
tuning ») et à la coordination de mouvements produits ailleurs dans
le cerveau, et intègre le tout pour produire des mouvements si fluides
et harmonieux que nous n’en avons même plus conscience.
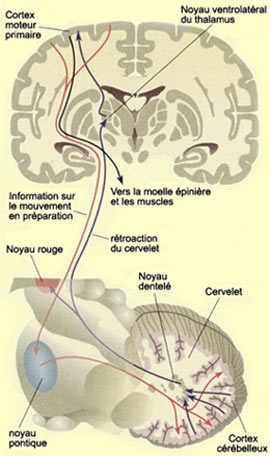
Pour ce faire, le cervelet entretient d’étroites
relations avec le cortex. Les aires motrices, somatosensorielles et pariétales
postérieures de ce dernier envoient un important contingent d’axones
vers les noyaux du pont situés dans le tronc cérébral. Les
neurones du pont projettent ensuite leurs axones dans le cervelet. Cette voie
corticopontocérébelleuse forme un faisceau d’une extrême
densité qui contient environ 20 millions d’axones, soit à
peu près 20 fois plus que le faisceau pyramidal !
Les deux hémisphères
cérébelleux projettent en retour vers le cortex moteur par une voie
impliquant le thalamus. Cette voie fait relais au niveau du noyau ventrolatéral
(VLc) du thalamus. Les hémisphères cérébelleux influencent
ainsi la musculature des membres via le cortex et le
système moteur latéral.
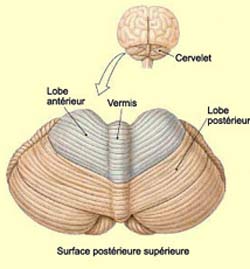
Les deux hémisphères
du cervelet ne sont pas divisés nettement en deux comme les deux hémisphères
du cerveau. La partie médiane constitue ce qu’on appelle le vermis
cérébelleux. Le vermis ne présente pas de latéralisation
et envoie des axones vers le tronc cérébral qui, par l’entremise
du système
ventromédian, contribue au maintien de la posture.
Les mécanismes cérébraux
derrière la préparation et l’exécution d’un mouvement
dépassent la vision simpliste du cortex moteur qui donne les ordres que
les motoneurones exécutent.
En effet, imaginez que vous voulez
prendre un verre d’eau fraîche qui est en réalité rempli
d’eau bouillante. Dès que vous touchez le verre, vous retirez immédiatement
la main de façon réflexe,
sans y penser.
Sachant maintenant que le contenu du verre est chaud,
vous pourriez tout de même le soustraire à votre enfant qui tente
de l’attraper grâce au contrôle volontaire du mouvement. Vous
contrez alors consciemment le réflexe de retrait et saisissez tout de même
le verre d’eau bouillante car l’enjeux est d’une grande importance
pour vous.
Enfin si l’on vous dit que le verre est en cristal
et non en verre. La façon dont vous le prendrez sera sans doute plus délicate.
Autrement dit, votre cerveau tiendra compte de cette information et adaptera sa
façon de saisir le verre en conséquence.
L’idée
que l’exécution motrice est le résultat d’une construction
très élaborée s’impose donc sur l’idée
que le contrôle moteur serait une simple commande qui dirait « go
» aux motoneurones de la moelle épinière. De plus, la remarquable
adaptabilité de l’activité motrice révèle l’intervention
de puissants mécanismes
de régulation et de rétroaction.
LA SÉQUENCE D'ACTIVATION DES AIRES MOTRICES
On peut distinguer trois
opérations dans le traitement de l’information nécessaire
au déclenchement d’un mouvement volontaire. La première consiste
à sélectionner une réponse adaptée
à la situation dans un répertoire de réponses possibles.
Cette réponse qui correspond à un objectif comportemental particulier
est déterminée de façon globale et symbolique.
La
deuxième opération en est une de planification concrète
du mouvement. Elle consiste à définir les caractéristiques
de la réponse sélectionnée en terme de séquences de
contractions musculaires nécessaires pour réaliser la réponse
choisie.
La troisième opération consiste à exécuter
le mouvement proprement dit. C’est elle qui est à l’origine
de l’activation des motoneurones responsables de la mécanique observable
du geste.
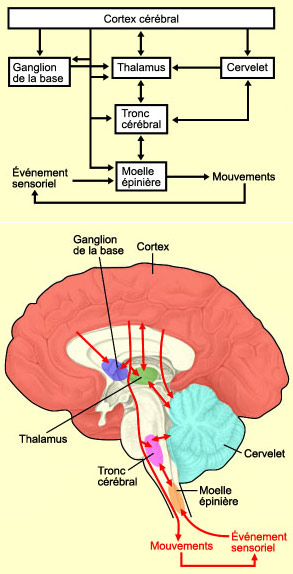
Par conséquent,
les messages de commande émis par le cortex moteur sont eux-mêmes
déclenchés par des messages provenant d’autres
aires corticales. Le cortex moteur est aussi en étroite relation avec
des structures sous-corticales comme les
ganglions de la base et le cervelet
qui communiquent avec le cortex via la structure de relais que constitue le thalamus.
À
la lumière de ce que l’on sait de la séquence d’activation
des aires motrices du cortex, on peut donc décortiquer la commande classique
« Attention… Prêt ? Partez ! » en terme d’activité
cérébrale localisée.
Les lobes pariétaux
et frontaux entrent d’abord en activité, avec une contribution des
structures sous-corticales impliquées dans la vigilance et l’attention.
Le « prêt? » active par la suite les aires corticales supplémentaires
et prémotrices où les stratégies du mouvement sont élaborées
et maintenues jusqu’au signal du départ. L’ordre « partez
! » peut venir de l’extérieur comme lors d’une course,
ou de la personne elle-même lorsqu’elle décide que les conditions
sont réunies pour déclencher son action. Cette commande fait alors
intervenir des informations issues de structures sous-corticales comme les ganglions
de la base qui vont influencer l’aire 6, puis éventuellement le cortex
primaire qui va réaliser l’action.