La rétine n’utilise
pas seulement la neurotransmission classique avec synapse
chimique pour transmettre l’information dans ses réseaux.
Beaucoup d’information se transmet en effet rapidement
par des synapses
électriques, surtout dans les voies partant des
bâtonnets. De plus, on a découvert aussi beaucoup
de neuromodulation dans
la rétine, c’est-à-dire de substances
diffusant à distance et influençant un grand
nombre de neurones à la fois.
Les cellules ganglionnaires sont
donc le dernier maillon de la chaîne neuronale rétinienne.
L’information recueillie par 125 millions de photorécepteurs
par oeil est canalisée dans un million de cellules
ganglionnaires.
Les potentiels d’action qui sortent de l’oeil par
le nerf optique après leur passage dans les quelques
synapses de cette chaîne sont donc porteurs d’une
information beaucoup plus sophistiquée que la représentation
point par point du monde encodée par les photorécepteurs.
La rétine n’est donc pas un simple feuillet de
cellules photosensibles, mais véritablement un petit
cerveau à l’extérieur du gros ! Elle se
détache d’ailleurs de celui-ci lors du développement
embryonnaire.
LA RÉTINE
À l’instar des cônes et des bâtonnets
dont la
structure et la
fonction sont toutes entières orientées vers la
transformation de l’énergie lumineuse en influx
nerveux, chaque
type de cellule de la rétine possède une localisation
et une connectivité visant à effectuer un premier traitement
de l’information.
Source : University of Kansas Medical Center
Alors que les autres
neurones de la rétine n’émettent que
des potentiels électriques gradués, les cellules
ganglionnaires sont les seules à transmettre
le signal nerveux sous forme de potentiels
d’action. Considérant que ce sont leurs
axones qui forment le nerf optique et transmettent donc l’information à de
grandes distances de la rétine, la génération
de potentiels d’action dans ces cellules prend alors
tout son sens. Ces potentiels d’action sont d’ailleurs
générés de façon spontanée
et c’est donc leur fréquence de décharge
qui est amplifiée ou diminuée par l’apparition
de lumière dans leur champ récepteur.
Bien que la plupart des cellules ganglionnaires ont soit
des champs récepteurs de type centre ON –
périphérie OFF, soit l’inverse,
il existe d’autres critères définissant
d’autres catégories. Leur aspect général,
leurs connexions et leurs caractéristiques électrophysiologiques
ont permis de distinguer au moins trois types de cellules
ganglionnaires dans la rétine des macaques, une
rétine très semblables à la nôtre.
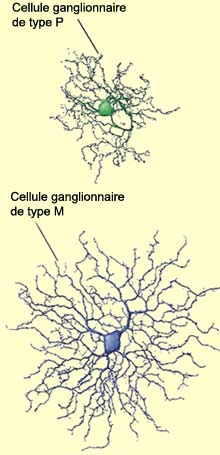
De petites cellules ganglionnaires de type
P (pour parvus,
petit en latin) qui représentent environ 90% de la population
totale de cellules ganglionnaires; de grandes cellules de type
M (pour magnus, grand en latin) qui constituent environ
5% de la population; et des cellules ganglionnaires non M-non P
qui ne sont pas encore bien caractérisées et qui
forment le 5% restant.
Les cellules de type
M présentent aussi de plus grands champs
récepteurs, propagent les potentiels d’action
plus rapidement dans le nerf optique, et sont plus sensibles
aux stimuli à faible contraste. De plus, la réponse
positive d’une cellule M à une stimulation est
une brève salve de potentiels d’action, alors
que les cellules P ont une réponse plus tonique et
maintenue aussi longtemps que le stimulus agit.
L’idée la plus couramment admise est que les cellules
M sont particulièrement impliquées dans la détection
du mouvement du stimulus alors que les cellules P, avec leur
petit champ récepteur, seraient plus sensibles à la
forme et aux détails de celui-ci.
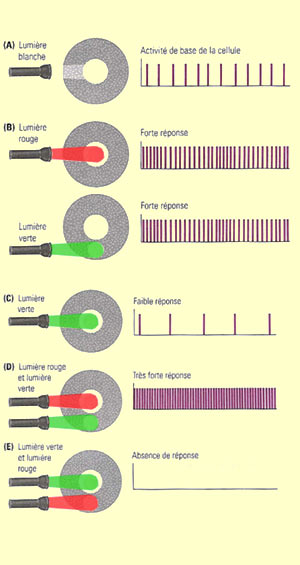
Autre distinction essentielle
pour la détection de la couleur : les cellules P et
quelques cellules non M-non P sont sensibles
aux différences de longueur d’onde de la lumière.
La majorité des cellules P sont en fait des « cellules à opposition
simple de couleur », c’est-à-dire que la
réponse à une longueur d’onde donnée
au centre de leur champ récepteur est inhibée
par la réponse de la périphérie à
une autre longueur d’onde. Dans le cas d’une cellule à centre
rouge ON et périphérie verte OFF, ce sont des
cônes rouges qui occupent le centre du champ et des verts
la périphérie. Même chose pour les cellules à opposition
bleu-jaune, où ce sont des cônes bleus versus
des cônes sensibles au rouge et au vert. Les cellules
ganglionnaires de type M n’ont pas d’opposition
à la couleur tout simplement parce qu’à
la fois le centre et la périphérie reçoivent
de l’information de plus d’un type de cône.
Les cellules de type M ne se retrouvent pas non plus dans la
fovéa ce qui confirme l’absence de rôle
dans le traitement de la couleur pour ces cellules.
Les cellules ganglionnaires apportent donc au cerveau de l’information
en partie déjà traitée en ce qui a trait à la
comparaison régionale des processus d’opposition lumière-obscurité,
rouge-vert et bleu-jaune
.
Comme dans plusieurs systèmes
cérébraux, le système visuel traite
l’information en parallèle.
Nos deux yeux apportent d’abord deux flux d’information
parallèles que notre cerveau va comparer pour avoir une
estimation de la profondeur d’un objet donné dans
une scène.
Il semble qu’il y ait également des flux d’information
indépendants sur la quantité
de lumière à chaque point de l’espace transmis
par les cellules ganglionnaires.
Les cellules ganglionnaires de type M peuvent aussi détecter
des contrastes subtils grâce à
leur vaste champs récepteurs et les cellules P, avec
leurs petits champs récepteurs, semblent mieux adaptées à la
discrimination de petits détails. Enfin, les cellules
P et non M-non P sont spécialisées dans le traitement
des couleurs.
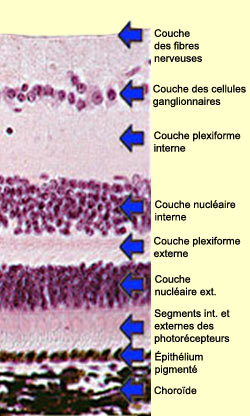
LES CHAMPS RÉCEPTEURS,
DE LA RÉTINE AU CORTEX
L’organisation
centre-périphérie des champs récepteur des
neurones rétiniens naît de la connectivité
des cellules horizontales avec les phororécepteurs
et les cellules bipolaires. Cette organisation permet d’augmenter
les contrastes des objets dans le champ visuel. Une rétroaction
en provenance de la couche
plexiforme interne influence aussi l’activité
des cellules horizontales. Celles-ci, en retour, modulent
les signaux des photorécepteurs sous différentes
conditions d’éclairage, rendant la
transduction moins sensible sous de fortes lumières
et plus sensible en lumière atténuée.
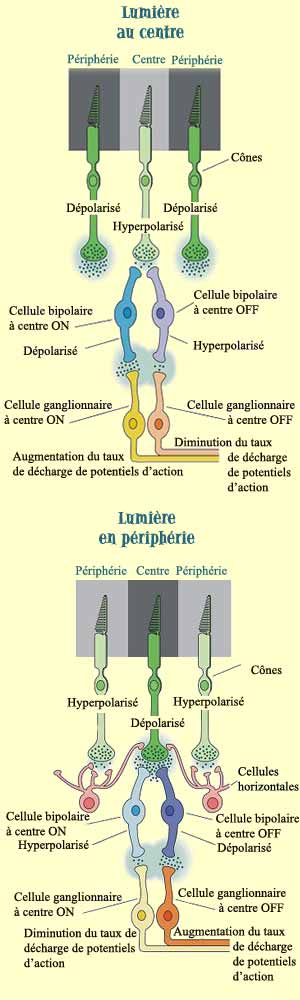
Les cellules bipolaires, comme tous les neurones de la rétine
sauf les cellules ganglionnaires, transmettent l’influx
nerveux non pas avec des potentiels d’action mais sous
la forme de simples potentiels gradués. On parle tout
de même de réponse ON lorsqu’une dépolarisation
amène une augmentation de la relâche de neurotransmetteurs
et de réponse OFF quand une hyperpolarisation diminue
la quantité
de neurotransmetteurs relâchés.
La question que l’on peut alors se poser est celle
de la nature, excitatrice ou inhibitrice, de la connexion
reliant les photorécepteurs aux cellules bipolaires.
Les photorécepteurs émettent en permanence leur
neurotransmetteur, du glutamate, en l’absence de lumière.
Par conséquent, les récepteurs
au glutamate des cellules bipolaires à centre OFF
sont excitateurs, puisque l’absence de lumière
doit les stimuler. De même, on constate que les récepteurs
des cellules bipolaires à centre ON sont inhibiteurs,
puisque la lumière qui frappe les photorécepteurs
au centre de leur champ récepteur va les hyperpolariser
et diminuer la relâche de glutamate. Moins de ce neurotransmetteur
qui est ici inhibiteur (dû au type de récepteur
métabotropique sur lequel il se fixe) amènera
donc une plus grande excitation du neurone bipolaire. C’est
donc la nature excitatrice ou inhibitrice des récepteurs
au glutamate qui détermine le type de champ récepteur
des cellules bipolaires.
Cellules ganglionnaires
à centre ON
Les cellules
ganglionnaires ont le même type de champs récepteurs
circulaire à opposition centre-périphérie
que les cellules bipolaires. De plus, le caractère ON
ou OFF d’une cellule bipolaire est transmis à
la cellule ganglionnaire qui lui est connectée. La plupart
des cellules ganglionnaires ne sont pas très sensibles
aux stimuli lumineux qui touchent à la fois le centre
et la périphérie du champ récepteur. Ainsi,
une obscurité totale ou un éclairage uniforme
leur fait émettre peu de potentiels d’action.
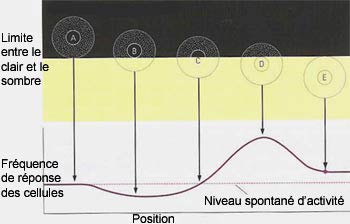
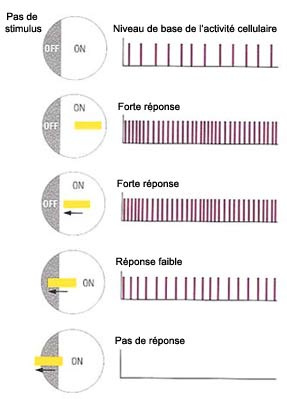
Ces cellules sont toutefois très sensibles aux différences
d’éclairement survenant à l’intérieur
du champ récepteur, comme lorsqu’une zone d’ombre
ou de lumière balaie leur champ récepteur d’un
côté
à l’autre par exemple.
L’information portée par les potentiels d’action
des cellules ganglionnaires souligne donc davantage les contrastes
d’éclairement entre les zones éclairées
et obscures plutôt que le degré absolu de luminosité. La
perception de la lumière et de l’obscurité n’est
donc pas absolue, mais relative.
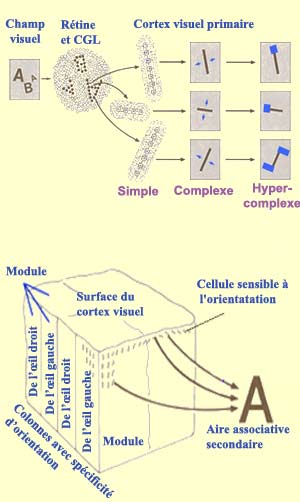
Outre les cellules simples et complexes de
l’aire visuelle primaire (aire 17 ou V1 ou encore
cortex strié) ou de l’aire secondaire (aire
18 ou V2), on trouve dans l'aire visuelle secondaire V5
ou (aire 19 ou MT) des cellules qui ne répondent
que si un stimulus lumineux présente un rapport
surface éclairée/surface sombre donné,
ou bien si un angle lumineux donné est présent,
ou encore à des formes en mouvement. Certaines de
ces cellules dites hypercomplexes ne sont également
sensibles qu’à
des lignes de longueur définie, de sorte que l'allongement
du stimulus au-delà de cette limite réduit
la réponse.
Les cellules hypercomplexes résultent de la convergence
sur un même neurone des axones de plusieurs cellules
complexes d'orientations différentes et de champs
visuels voisins. Elles constituent encore un autre niveau
de traitement de l'information. A chacun de ces niveaux,
chaque cellule «voit» plus que celles de niveau
inférieur et les cellules les plus hautes ont le pouvoir
d'abstraction le plus élevé. Ce dernier est
généré par la connectivité
neuronale à chaque
relais des voies visuelles jusqu’aux différents
cortex visuels.
On peut résumer ceux-ci de la façon suivante
: la rétine et le CGL « voient »
la position, les cellules simples l’axe d’orientation,
les cellules complexes le mouvement de l’axe et les
cellules hypercomplexes les bords et les angles.
En plus de sortir du cortex visuel
primaire, les axones des cellules pyramidales de toutes les
couches se ramifient et forment des connexions locales entre
elles. La plupart de ces connexions se font perpendiculairement à la
surface corticale, passant ainsi à travers les différentes
couches. En demeurant dans la même colonne, ces connexions
radiales préservent ainsi la rétinotopie.
Les axones de certaines cellules pyramidales de la couche III
se ramifient également mais forment plutôt leurs
connexions horizontalement, à l’intérieur
même de la couche III. Ces connexions radiales
et horizontales jouent des rôles distincts dans
l’analyse de l’information visuelle.
Dans le système
visuel, les afférences qui véhiculent l'information
venant des deux yeux convergent initialement sur les mêmes
cellules cibles. Quelques semaines après la naissance
cependant, une ségrégation survient et les connexions
se font alors en fonction de l'origine oculaire de l'influx. Chaque
couche du corps genouillé latéral ou chaque
colonne de dominance oculaire du cortex strié reçoit
alors des afférences d'un seul œil suite à
la réorganisation synaptique.
Plusieurs expériences visant à étudier
les effets de la privation sensorielle durant des périodes
critiques du développement ont été réalisées
chez le chat et le singe en suturant une ou deux paupières,
ou encore en créant chirurgicalement un strabisme
artificiel. Il ressort de ces études que le développement
normal de la connectivité du cortex
visuel ne dépend pas tant de l'activité
absolue d'une voie nerveuse particulière, mais bien
d'une compétition entre l'activité relative
de différentes voies.
Après que l'œil droit d'un jeune chat ait été
suturé durant la période critique de l'établissement
des colonnes de dominance oculaire dans le cortex visuel primaire,
un processus de compétition fait en sorte que la superficie
des colonnes innervée par les voies visuelles de l'œil
suturé diminue par rapport à celle de l'œil
intacte. Il semble que les cellules du corps genouillé
latéral recevant des connexions de l'œil fermé
voient dans un premier temps leur axone allant au cortex régresser.
Les neurones corticaux ainsi laissés vacants se voient
dans un deuxième temps innervés par des collatérales
s'étant développées à partir des
axones des cellules du corps géniculé latéral
de l'œil intact.
L'ORGANISATION
CELLULAIRE DU CORTEX VISUEL
Le grand nombre de travaux sur le cortex
visuel primaire nous permet d’appréhender
toute la complexité et la beauté
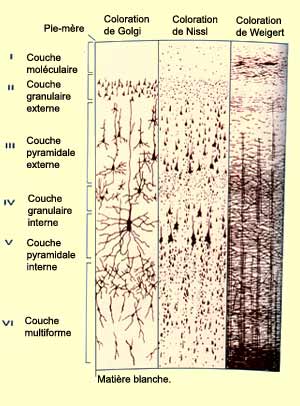
de son organisation cytoarchitecturale. D’abord une stratification
horizontale en différents types de neurones spécialisés
dans la réception ou l’envoi de l’information
nerveuse.
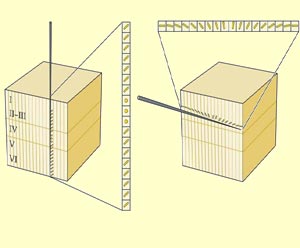
Ensuite une autre fragmentation, radiale celle-là,
en une multitude de colonnes où les
neurones qui s’y trouvent répondent tous à une
même caractéristique d’un point donné
du champ visuel. Les colonnes forment donc une unité fonctionnelle
perpendiculaire à
la surface corticale.
D’ailleurs, si on
introduit une microélectrode perpendiculairement
à travers les différentes couches du cortex visuel,
on ne rencontrera par exemple que des neurones qui ont la même
préférence d’orientation, qu’ils
aient des champs
récepteurs simples ou complexes. Le corollaire
étant qu’une microélectrode pénétrant
parallèlement à
la surface du cortex, donc progressant dans une seule couche
corticale mais traversant plusieurs colonnes, va voir la préférence
d’orientation changer au fur et à mesure qu’elle
avance. Hubel et Wiesel ont montré que la préférence
d’orientation était ainsi inversée de 180
degrés en moyenne lorsque l’électrode se
déplace de 1 millimètre environ dans la couche
III.
On peut dire que les colonnes
de dominance oculaire constituent un troisième
niveau d’organisation cellulaire dans le cortex visuel
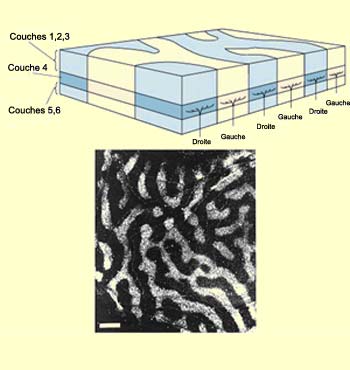
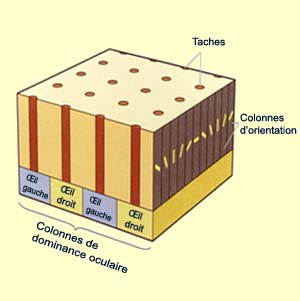
primaire. Ces colonnes, situées au niveau de la couche
IV C, se présentent en fait sous forme de bandes de
0,5 millimètres de large régulièrement
espacées. En fait, des expériences de marquage
(voir capsule expérience intermédiaire à
gauche) permirent de constater que ces bandes correspondaient
aux terminaisons nerveuses de l’œil gauche et de
l’œil droit et qu’elles alternaient ainsi
d’un œil à l’autre, se disposant un
peu comme les rayures d’un zèbre.
Comme si ce n’était
pas suffisant, d’autres chercheurs ont mis en évidence à la
fin des années 1970, à l’aide d’un
colorant appelé cytochrome oxydase, un autre type de
colonnes localisées à intervalles réguliers
et traversant les couches II, III, V et VI. Ces colonnes qui
ont l’aspect des taches de la robe d’un léopard
lorsque vues tangentiellement reçurent donc le nom de taches.
Ces taches sont donc disposées en lignes et centrées
sur une bande de dominance oculaire de la couche IV C. Entre
les taches se trouvent forcément des zones intermédiaires
dites « intertaches
» dont les neurones ne possèdent pas les caractéristiques
de ceux des taches.
Les cellules des taches ont ceci de particulier qu’elles
sont sensibles à la longueur d’onde du stimulus,
autrement dit à
sa couleur. De plus, elles sont monoculaires, n’ont pas
de spécificité d’orientation et présentent
plutôt des champs récepteurs
à symétrie circulaire. Dans certaines d’entre
elles, on retrouve même l’organisation centre-périphérie à opposition
de couleur des cellules ganglionnaires de types P qui sont à l’origine
de cette voie (voir encadré ci-bas).
Hubel
et Wiesel ont également montré
que chaque point du champ visuel produit une réponse
dans une région de 2 x 2 millimètres dans le
cortex. Or une telle surface corticale peut contenir deux groupes
complets de colonnes de dominance oculaire, 16 taches et des
zones intertaches pouvant contenir plus de deux fois toutes
les orientations possible sur 180 degrés. Ils appelèrent hypercolonne (ou
plus généralement module cortical) cette
région du cortex qui semble à
la fois nécessaire et suffisante à
l’analyse de l’image d’un point dans l’espace
visuel. Comme le cortex est une couche cellulaire continue
et qu'il est très difficile d'établir concrètement
les frontières de tels modules, l'existence de ceux-ci
d'un point de vue fonctionnel est encore débattue.
Au début des années
soixante, David Hubel et Torsten Wiesel (Prix Nobel de médecine
1981) furent les premiers à explorer les champs récepteurs
des neurones du corps genouillé latéral et
du cortex visuel à l’aide de microélectrodes.
Ils ont d’abord montré que les neurones du corps
genouillé latéral se comportaient pratiquement
comme les cellules ganglionnaires de la rétine. Puis,
ils ont mis en évidence trois voies relativement indépendantes
dans le traitement de l’information visuelle qui prennent
en charge chacune un aspect différent de la vision.
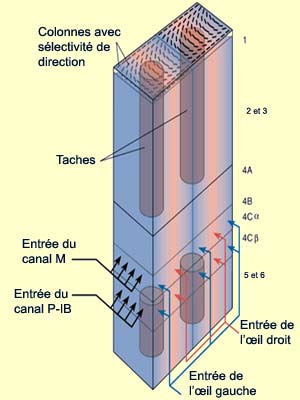
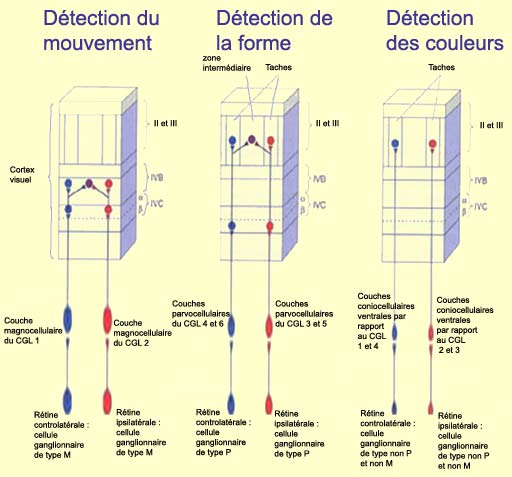
Le premier est le canal M pour «
magnocellulaire » qui commence avec les
cellules ganglionnaires magnocellulaires de la rétine,
fait un relais dans le corps
genouillé latéral et rejoint la couche IV
C alpha du cortex strié. À ce niveau, les champs
récepteurs des neurones de la couche IV C alpha ne sont
plus circulaires (comme c’était le cas dans la
rétine et dans le CGL), mais sont
plutôt allongés.
Les cellules de la couche IV C alpha projettent aux neurones
de la couche IV B qui ont eux aussi des champs récepteurs
simples mais répondent souvent
à des stimulations des deux yeux contrairement aux cellules
de la couche IV C dont les champs récepteurs sont monoculaires.
Les neurones de la couche IV B commencent donc le travail d’intégration
nécessaire
à notre vision binoculaire.
Les cellules de la couche IV B sont aussi sélectives
à la direction, mais à condition que le stimulus
rectiligne se déplace dans une direction précise.
Pour cette raison, le canal M est considéré
comme spécialisé dans l’analyse du déplacement
des objets.
La seconde voie de transmission de l’information visuelle
est le canal P-IB pour «parvocellulaires-intertaches»
(ou interblob, en anglais), qui débute dans
la rétine avec les
cellules ganglionnaires parvocellulaires, passe par
le CGL et aboutit sur les cellules de la couche IV C bêta
qui réagissent comme les cellules du CGL parvocellulaires
d’où elles proviennent : elles présentent
des petits champs récepteurs monoculaires de configuration
circulaires dont la plupart sont à opposition centre-périphérie
rouge-vert.
Les axones des cellules de la couche IV C bêta rejoignent
ensuite les régions intermédiaires situées
entre les zones de tache de la couche III. Les
cellules complexes qui s’y trouvent, encore plus
spécifiques que les cellules simples pour l’orientation
des stimuli, portent à croire que l’analyse de
la forme des objets une spécialité
du canal P-IB.
La troisième voie de transmission du signal visuel est
le canal des taches, qui passe
à travers les couches parvocellulaires et coniocellulaires
du CGL et converge ensuite sur les zones de tache de la couche
III. Outre certains neurones de la couche IV C, les cellules
des taches contiennent les seuls neurones sensibles à la
couleur du cortex strié
et ces zones de tache servent donc probablement à
l’analyse de la couleur des objets.