Richesses
et limites
du concept de « modularité
cérébrale »
d’un point de vue neurobiologique
I-
En guise d’intro : l’outil « Le cerveau à tous les niveaux »
II-
Zoom in et zoom out à l’intérieur du cerveau humain
III-
Voyage dans le temps : la morphologie
évolutive des chordés
IV-
Richesses et limites
de la psychologie évolutive
1)
Les « sept péchés capitaux » de la psychologie évolutives pour un neurobiologiste
2)
Où sont les modules ?
--
I- En guise d’intro : l’outil « Le cerveau
à tous les niveaux »
Il
y a 5-6 ans, j’ai conçu le site le cerveau à tous les niveaux
(www.lecerveau.mcgill.ca ) avec
lequel on va travailler aujourd’hui, et que je présente souvent comme le fruit
des frustrations accumulées au cours des 10 années où j’ai travaillé en communication
scientifique sur le cerveau. Frustrations qui j’ai tenté de résoudre par la
structure même du site, i.e. un site où tout est expliqué à la fois en terme
simple pour les débutants (cliquer
sur un thème), mais aussi de plus en plus compliqué pour les intermédiaires (cliquer) et les avancés (cliquer, remarquer code de couleur);
et aussi un site où chaque sujet est
systématiquement exploré à 5 niveaux
d’organisation, du moléculaire (cliquer) au cellulaire (cliquer), au cérébral
(cliquer), au psychologique (cliquer), et au social (cliquer), tout simplement
parce qu’avec le cerveau, à chaque fois qu’il se passe quelque chose à un
niveau, il se passe toujours, en même temps autre chose à tout les autres
niveaux et une compréhension globale du comportement humain implique pour
moi que l’on prennent en compte l’ensemble de ces phénomènes dans leur simultanéité.
En
même temps, j’ai toujours cru qu’une véritable
compréhension de l’anatomie et du fonctionnement du cerveau, et par conséquent
des comportements humains, nécessitait d’avoir constamment à l’esprit une
perspective évolutive. D’où un thème au complet parmi les 12 de la
page d’accueil intitulé le Bricolage de l’évolution que j’ai consacré à l’évolution,
contrairement à bien des bouquins sur le cerveau qui se contentent de rappeler
en quelques pages au début de l’ouvrage que l’on a une longue histoire évolutive
mais sans vraiment en faire sentir l’énorme poids et les innombrables conséquences.
Comme
je m’adresse à des étudiants en philosophie qui n’ont peut-être pas à l’esprit
les principaux repères d’anatomie cérébrale, je vous ai préparé une
petite visite à travers le site pour vous rafraîchir la mémoire sur les
différentes structures cérébrales. Cela, dans une perspective évolutive et
dans le but, je l’espère, de permettre d’ouvrir ensuite la discussion sur
le sujet pour lequel vous m’avez invité, c’est-à-dire la « modularité
cérébrale ».
II- Zoom in et zoom out à l’intérieur du cerveau
humain
On
va donc partir du cerveau et descendre progressivement aux neurones, puis
aux synapses et finalement aux molécules. Ensuite, on va revenir sur nos pas
en faisant un espèce de zoom out,
où l’on va remonter jusqu’au niveau cérébral mais en ayant à l’esprit peut-être
un petit peu plus ce que ce cerveau contient et la complexité de ses processus.
On
commence par considérer le cerveau humain au
niveau macroscopique, ce qui correspond dans le site au niveau « cérébral »
dans le site (https://lecerveau.mcgill.ca/flash/a/a_01/a_01_cr/a_01_cr_ana/a_01_cr_ana.html
)
On
a ici en haut un cerveau humain vu de l’extérieur, et en dessous une coupe sagittale médiane (i.e. qu’on a
coupé sur le sens de la longueur exactement entre les deux hémisphères et
on regarde à l’intérieur)
Je
vais vous présenter rapidement la nomenclature qu’utilisent les neurobiologiste
pour classifier les différentes structures macroscopiques du cerveau parce
qu’elle est inspirée de l’origine évolutive
de ces structures et qu’on va y revenir plusieurs fois durant la présentation.
La
première chose à dire, en fait, c’est qu’il y a dans le cerveau des parties
plus anciennes que d’autres, et qu’on peut grosso modo établir un gradient
d’ancienneté de la partie postérieure à la partie antérieure. Autrement dit,
ce qu’on appelle le rhombencéphale
est la partie la plus ancienne du cerveau. On y distingue le myélencéphale (ex.) et le métencéphale
(ex.). On distingue ensuite le mésencéphale,
qui est donc une structure un peu plus récente (ex.). Et finalement le prosencéphale,
la partie la plus récente du cerveau, qu’on subdivise en diencéphale (ex.) et en télencéphale
(ex.) qui comprend, entre autre, le fameux cortex cérébral qui est très développé
chez l’humain comme vous le voyez.
(puis descendre
au bas de la page)
Évidemment
ce cerveau n’est pas séparé du reste du corps, et on n’insistera jamais assez
pour dire que ce n’est pas le cerveau qui commande le corps, mais bien que
les deux sont liés de manière inextricable, s’informant
mutuellement de manière constante, le corps envoyant autant de signaux qui
influencent le cerveau que le cerveau envoyant des signaux qui influencent
le corps.
Cela
se fait évidemment par la moelle épinière qui est le prolongement naturel
du bulbe rachidien (que l’on a vu tantôt) dans la colonne vertébrale et qui
forme avec lui ce qu’on appelle LE
SYSTÈME NERVEUX CENTRAL. L’autre grande partie de notre système nerveux, c’est
ce qu’on appelle LE SYSTÈME NERVEUX PÉRIPHÉRIQUE i.e. les nerfs qui parcourent tout le corps, dont certains recueillent
de l'information et d'autres diffusent des ordres.
Ce
système nerveux périphérique, on le subdivise en système nerveux somatique et en système nerveux végétatif. (là c’est un peu didactique mon
affaire, mais ça sera pas long, c’est juste pour être sûr qu’on a un portrait
global de l’affaire…)
Donc
un certain nombre de ces neurones et de leurs prolongements qui forment des
nerfs, c’est le système nerveux somatique
qui participent à la vie de relation de l'organisme avec son milieu extérieur. Ils transmettent au
cerveau de l'information en provenance des différents détecteurs sensoriels.
Et ils nous permettent de répondre à ces stimulations en bougeant dans cet
environnement
D’autres
neurones et leurs prolongements sont appelés le système nerveux végétatif, qui est celui qui intervient plutôt dans la régulation des fonctions vitales internes.
Ils contribuent à l'équilibre de notre milieu intérieur en coordonnant des
activités comme la digestion, la respiration, la circulation sanguine, l'excrétion
ou la sécrétion d'hormones.
Ce
système nerveux végétatif est à son tour subdivisé en
SYSTÈME NERVEUX SYMPATHIQUE dont l’activation
prépare l'organisme à l'activité physique ou intellectuelle. Et en SYSTÈME NERVEUX PARASYMPATHIQUE dont
l’activation amène un ralentissement
général des fonctions de l'organisme afin de conserver l'énergie.
--
Maintenant, un mot sur les cellules
nerveuses et leur mode de communication. On va donc quitter le niveau
cérébral pour descendre au niveau neuronal, au niveau des cellules nerveuses
qui forment ce système nerveux. Je vous rappelle ici vraiment juste le minimum,
toujours dans le but de se rappeler de quoi on va parler concrètement quand
on va aborder la question de la modularité cérébrale…
J’ai trouvé un site bien intéressant que j’aimerais vous montrer pour faire
cette transition du niveau cérébral au niveau neuronal. C’est en fait le premier
effet spécial « hight tech » d’une longue série que vous aller avoir la chance
de voir au cours de cette présentation…
http://brainmaps.org/ :
cliquer sur le « sample » de coupe de
cerveau de primate (avec zoom in jusqu’au niveau des neurones)
(revenir à mon
site, remonter la page et cliquez sur cellulaire)
Voici
donc une photo de quelques cellules
nerveuses avec leur prolongements
plus visibles et avec une micro-électrode
comme on s’en sert pour faire des enregistrements intracellulaires de l’activité
électrochimique des neurones.
(cliquer https://lecerveau.mcgill.ca/flash/d/d_01/d_01_cl/d_01_cl_ana/d_01_cl_ana.html
)
Les
neurones sont des cellules du corps
humain spécialisées dans la communication, et cette spécialisation détermine
leur forme. À partir du corps cellulaire se forme donc 2 types de prolongement,
les dendrites qui sont habituellement plus courts et multiples, et dont la
fonction est grosso modo de recevoir l’information, et l’axone, généralement
unique et très long, qui envoi l’influx nerveux vers d’autres neurones.
(cliquer https://lecerveau.mcgill.ca/flash/d/d_01/d_01_cl/d_01_cl_fon/d_01_cl_fon.html)
C’est
ainsi que les neurones peuvent former des circuits où l’information est transmise d’un neurone à l’autre.
(cliquer sur https://lecerveau.mcgill.ca/flash/i/i_01/i_01_cl/i_01_cl_ana/i_01_cl_ana.html
)
Si
on agrandit maintenant la connexion entre deux neurones, on constate qu’il
ne se touchent pas, mais que les deux membranes sont séparées par un très
mince espace, et c’est ce qu’on appelle la
synapse, dont le fonctionnement est à la base de tous nos processus mentaux.
Je
vais y revenir à l’instant à la synapse, mais je veux juste vous mentionner
avant l’existence d’un autre type cellulaire dans le cerveau, soit les cellules gliales, qui servent entre autre d’isolant aux axones, ce
qui permet d’accélérer la propagation de l’influx nerveux, et aussi qui remplissent
toutes sortes d’autres fonctions complexes de nutrition mais aussi de communication,
des fonctions encore mal connues. Et considérant qu’il y a environ 100
milliards de neurones dans le cerveau, mais qu’il y a environ 10
fois plus de cellules gliales dont on entend cependant encore très peu
parler, cela donne une idée de la complexité de l’objet dont on parle.
Très
rapidement toujours, on va maintenant faire un soupçon de physiologie nerveuse en allant ici (cliquez sur
https://lecerveau.mcgill.ca/flash/d/d_01/d_01_m/d_01_m_fon/d_01_m_fon.html
)
ou
l’on voit la région du corps cellulaire d’un neurone d’où part l’axone, simplement
pour montrer qu’il y a une différence
de charge électro-chimique (des ions, i.e. des
sels comme sodium, potassium…) entre l’intérieur (qui est plus négatif) et
l’extérieur de la cellule (qui est plus positif) et que ce qu’on appelle un
potentiel d’action, ou un influx nerveux,
c’est simplement une inversion rapide et transitoire de ce potentiel de membrane
qui se déplace le long de l’axone.
(cliquez sur 2, puis sur 3)
(allez à https://lecerveau.mcgill.ca/flash/i/i_01/i_01_cl/i_01_cl_fon/i_01_cl_fon.html
)
ici on
voit 3 neurones converger vers un 4e. Actuellement le neurone 1
émet spontanément des potentiels d’action
qui sont, comme on vient de le voir, des rapides changements de polarité de
la membrane.
Deux
choses importantes ici : 1) que le langage, si vous voulez, par lequel
les neurones communiquent entre eux est
la fréquence de ces potentiels d’action. On dit par exemple qu’un neurone
est excité ou qu’il répond à un stimulus lorsqu’il émet de nombreux potentiels
d’action; c’est par la fréquence
des potentiels d’action que l’information peut être transmise dans le cerveau;
2) que la plupart des neurones ont une activité
spontanée comme celui-ci et que c’est donc par la modulation de cette activité spontanée que l’information est
transmise.
Mentionnons
aussi que les P.A. sont des phénomènes tout
ou rien d’amplitude constante (il n’y a pas de « demi P.A.), contrairement
à se qui se passe de l’autre côté de
Cette
caractéristique des potentiels excitateur va être à la base de l’activité
computationnelle des neurones puisqu’elle va permettre la sommation temporelle de différents
influx nerveux. Par exemple on voit ici que le potentiel excitateur généré
par la synapse du premier neurone se
résorbe avant d’avoir atteint la base de l’axone d’où partent les potentiels
d’action. Mais si par exemple 2 potentiels d’action arrivent en même temps
sur le neurone (cliquez sur 2),
il y a sommation des deux potentiels excitateur et cette fois la résultante
a assez d’intensité pour atteindre le seuil de déclenchement d’un
nouveau P.A. qui part dans l’axone du second neurone vers d’autres neurones.
De la même façon, (cliquez sur 3),
il peut y avoir des potentiel inhibiteur
qui peuvent contrecarrer l’effet des premier et ramener sous le seuil de déclenchement
du P.A. le potentiel post-synaptique, empêchant ainsi le P.A. de se propager.
Voilà
donc décrit en très grands traits les bases de la communication neuronale.
Je
vais maintenant faire une brève, mais alors là très brève incursion au niveau
moléculaire pour parler du fonctionnement
de la synapse parce qu’on ne peut passer à côté de cet élément à la base
de la plasticité synaptique, donc de l’apprentissage, de la mémoire et d’une
foule d’autres phénomènes comme vous allez le constater…
(aller à https://lecerveau.mcgill.ca/flash/i/i_07/i_07_m/i_07_m_tra/i_07_m_tra.html)
Donc
on a ici une synapse, avec le bout de l’axone d’un premier neurone pré-synaptique
en haut, et le bout d’une dendrite d’un second neurone post-synaptique en
bas ici. À l’intérieur de la terminaison axonale,
on retrouve plusieurs vésicules
contenant des molécules qu’on appelle des neurotransmetteurs,
qui sont comme des clés qui vont aller se fixer sur des récepteurs qui sont d’autre plus grosses molécules, des protéines
en fait, qui sont fichées à travers
Maintenant,
j’ai mentionné tantôt que les synapses était importantes pour la plasticité
cérébrale, i.e. pour l’apprentissage. C’est en effet en modifiant l’efficacité synaptique que l’on va renforcer ou affaiblir
des circuits nerveux, et donc emmagasiner des apprentissages dans notre mémoire.
Je
vais vous montrer ici une façon par laquelle un neurone peut renforcer sa
connexion avec un autre. Il s’agit d’un processus appelé « potentialisation
à long terme » qui a été l’un des premier à être identifier, vers
les années 1970 si je ne m’abuse. La PLT survient lorsque le premier neurone
est activé par de nombreux potentiel d’action (cliquez
sur bouton 2 de l’anim, ce qui a pour effet
de faire sortir plus de neurotransmetteur, et donc d’activer plus de récepteur,
et finalement de dépolariser davantage le second neurone, suffisamment pour
activer un second type de récepteur
en enlevant un espèce de bouchon constitué d’un atome de magnésium qui l’obstruait.
Or cet autre type de canal laisse entrer du Cacium en plus d’un peu de Sodium.
Et ce calcium va avoir plusieurs effets dans le neurone.
(remontrer l’animation)
(Cliquer
sur https://lecerveau.mcgill.ca/flash/a/a_07/a_07_m/a_07_m_tra/a_07_m_tra.html)
Et
là se complique un peu, comme vous le voyez, mais je ne vais pas rentrer dans
les détails, juste vous dire que le calcium, que l’on retrouve ici en orange, va activer certains enzymes, dont certains vont avoir un effet
facilitateur sur les récepteurs canaux déjà existants (i.e. ils vont s’ouvrir
plus longtemps par exemple), et d’autres vont remonter jusqu’au noyau du neurone dans le corps cellulaire, pour
dire aux gènes qui s’y trouve de
carrément construire de nouveaux récepteurs, qui vont par la suite être acheminés
jusqu’à la synapse au bon endroit pour encore une fois faciliter le passage
de l’influx nerveux.
Et
donc à ceux qui demande quelle est la
trace concrète matérielle de nos souvenirs, on peut dire, grosso modo,
que ce sont des récepteurs membranaires phosphorylés
(ou sensibilisé), et de nouveaux récepteurs synaptiques, donc des protéines
transmembranaires situés à sur des synapses spécifiques.
Mais
ces traces protéiques, seules, ne seraient rien si elles n’étaient comprise
dans un réseau de plusieurs centaines,
voir plusieurs dizaines de milliers de synapses qui relient autant de neurones
particuliers.
On
donc va maintenant commencer notre zoom
out et remonter au niveau cellulaire
(cliquer sur https://lecerveau.mcgill.ca/flash/d/d_07/d_07_cl/d_07_cl_tra/d_07_cl_tra.html
)
pour donner
une idée de la formation de certains
circuits facilités, ou d’assemblées de neurones, qui peuvent se former
dans le cerveau suite au renforcement de certaines synapses. Ces assemblées
neuronales peuvent donc former de véritables cartes mentales, tantôt d’une perception, tantôt d’une intention ou
d’un concept plus abstrait. Certains neurones peuvent participer simultanément à plusieurs de ces assemblées
neuronales, ce qui pourrait par exemple expliquer le caractère si associatif
de notre mémoire, le fait qu’une idée en appelle une autre, puis une autre,
etc. ce qui pourrait correspondre à une espèce de contagion d’une assemblée
de neurones à une autre…
Là
je viens de vous parler de circuits neuronaux en mettant l’emphase sur leur
plasticité, mais il y a aussi dans le cerveau humain d’innombrables circuits
qu’on pourrait qualifier de « précâblés », et dont la précision du câblage
dépend d’une part des gènes et
d’autre part de l’interaction avec
l’environnement durant le développement.
Le développement du cerveau étant un sujet fascinant que je mets pour l’instant
de côté pour garder une certaine suite dans mes idées, mais sur lequel j’espère
on aura le temps de revenir un peu plus tard…
(cliquer sur https://lecerveau.mcgill.ca/flash/a/a_01/a_01_cr/a_01_cr_fon/a_01_cr_fon.html
)
On
en voit ici quelques uns de ces circuits qui sont si importants qu’on voit
les faisceaux d’axones même à l’œil nu dans
une coupe de cerveau.
(descendre un
peu)
Plusieurs
autres circuits ont aussi été décrits au niveau
microscopique par les neurophysiologistes.
On a représenté ici par exemple qu’une partie de ceux qu’on connaît dans le
système visuel…
Il
y a donc dans le cerveau un circuiterie
précise qui permet d’accomplir certaines fonctions, et dont le niveau
de plasticité peut varier.
Mais
il y a aussi d’innombrables neurones qui fonctionnent selon une autre logique
si l’on veut, des neurone qui vont plutôt moduler les connexions de ces circuits en déversant dans de vastes
régions corticales des substances neuromodulatrices,
qui vont élever ou abaisser le seuil
d’excitabilité de vastes populations neuronales.
(descendre)
Ces
neurones sont généralement situés dans des amas de cellules du tronc cérébral et vont projeter des substances comme la noradrénaline, la sérotonine, l’acétylcholine
ou la dopamine dans différentes régions du cortex. Ce cerveau « neuro-hormonal »
comme on l’appelle souvent est la contrepartie du cerveau câblé, le Mr. Hyde du Dr. Jeckyll si vous voulez,
ou pour reprendre des classiques que vous connaissez peut-être, le Biologie
des passions de Jean-Didier Vincent par rapport à L’homme neuronal de Jean-Pierre
Changeux… Ce cerveau hormonal va donc être beaucoup associé à des états globaux
du cerveau comme les émotions, l’éveil, le sommeil, etc…
----
Voilà
donc ce qui va clore cette très rapide petite visite guidée et qui va me permettre
de faire une transition vers des considérations
plus évolutives concernant le cerveau.
(cliquer sur https://lecerveau.mcgill.ca/flash/d/d_05/d_05_cr/d_05_cr_her/d_05_cr_her.html)
Je
viens de parler un peu du tronc cérébral avec le cerveau hormonal, et aussi
des émotions. Parce que ce qu’il ne faut pas oublier c’est qu’en plus de ses
facultés cognitives dites « supérieures » qui permettent le langage
et la philosophie par exemple, l’être humain continue d’avoir des pulsions,
des besoins fondamentaux, des passions et des émotions. Nous sommes donc un amalgame de structures cérébrales récentes
avec d’autres plus anciennes et conservées au cours de l’évolution.
La
façon la plus synthétique de concevoir la structure
du cerveau en relation avec son histoire nous vient de Paul MacLean et de son fameux « cerveau triunique » proposé en 1970. Il
s’agit bien sûr d’une généralisation très approximative, mais elle constitue
néanmoins une bonne porte d’entrée à une perspective évolutive du cerveau.
Pour MacLean donc, trois
cerveaux distincts apparus successivement au cours de l’évolution cohabitent
en nous :
(cliquez sur les
3)
|
un
cerveau « reptilien », le plus ancien, qui assure
les fonctions vitales de
l’organisme, la fréquence cardiaque, la respiration, etc. qui comprend
le tronc cérébral et le cervelet, essentiellement
ce qui forme le cerveau d’un reptile. Il est fiable mais plutôt rigide. |
un
cerveau « limbique
», apparu avec les premiers mammifères, capable de mémoriser les comportements agréables ou désagréables, et par conséquent
responsable chez l’humain de ce que nous appelons les émotions. Il comprend principalement
l’hippocampe, l’amygdale et l’hypothalamus.
|
un
« néo-cortex », qui prend de l’importance
chez les primates et culmine chez l’humain avec nos deux gros hémisphères cérébraux. C’est grâce à
eux que se développera le langage,
la pensée abstraite, l’imagination, |
Or
ces trois cerveaux ne fonctionnent pas de manière indépendante et ont tissé de nombreuses connexions par lesquelles ils peuvent s’influencer
mutuellement. Les voies nerveuses qui vont du système limbique au cortex sont
par exemple particulièrement développées.
Le
modèle de MacLean a aussi la vertu de nous rappeler que les autres mammifères
et les reptiles sont nos ancêtres et que nous avons bel et bien hérité de certaines structures cérébrales
que possédaient ces animaux, comme nous avons hérité de leur structure corporelle bilatérale ou de
leurs quatre membres.
Ce
qui nous amène à la 3e partie de la présentation, qui s’intitule :
III- Voyage dans le temps : la morphologie évolutive
des chordés
Là
j’ai préparé quelque chose pour tenter de nous « faire sentir » un peu la durée de
l’évolution, mais je ne suis pas
trop sûr de mon coup alors si je divague un peu trop ou si tout ça est évident
pour vous dites-le moi et je passerai à la section suivante…
L’idée
de cette démonstration m’est donc venue de la très grande difficulté à s’extraire
de l’échelle temporelle humaine d’une centaine
d’année dans laquelle se déroule une vie humaine, pour envisager les millions d’années durant lesquelles a
eu lieu son évolution. Et aussi parce que je pense qu’ayant ressenti un
peu mieux ces durées géologiques, on pourra réfléchir peut-être mieux sur
la façon dont les circuits spécialisés du cerveau ont pu être façonnés par
l’évolution…
Pour
se représenter plus facilement ces durées extraordinaires, il est pratique
de ramener l’échelle des temps géologiques sur une horloge de 12 heures.
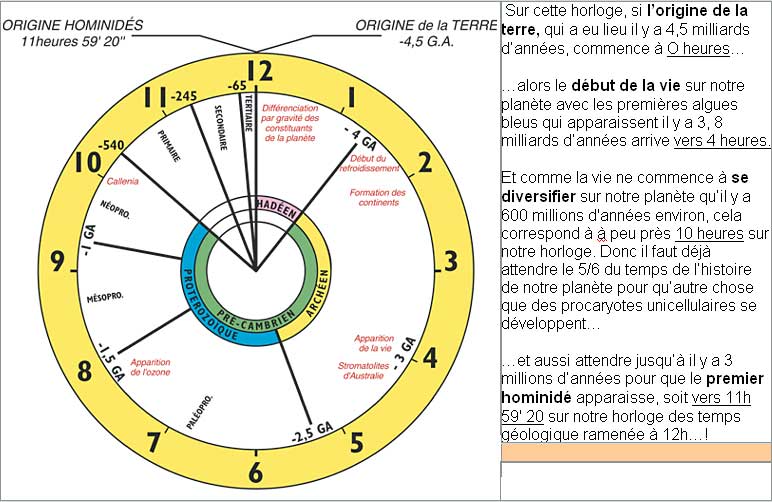
(Source : http://www.3d-fossiles.com/siences_de_la_vie/paleontologie/echelle_temps_geologiques_2.html
)
Mais si les premiers organismes pluricellulaires dotés d’un système nerveux sont apparus autour de 600 millions d’années avant aujourd’hui, et que c’est sur ce système nerveux que l’on veut réfléchir aujourd’hui, j’ai pensé qu’il serait utile de se représenter un peu mieux la durée de l’histoire de l’évolution des systèmes nerveux chez les animaux depuis son origine autour d’il y a 600 millions d’années. Et c’est ici que vous aurez droit à toute la technologie du XIXe siècle pour tenter de vous faire apprécier cette durée… Je vais donc sortir mon matériel à effet spéciaux ultra hight-tech !
[sortir la première bande
]
Ça
faisait longtemps que je voulais essayer ça et comme que vous m’en avez un
peu donné l’occasion, alors je me suis fait plaisir…]
Il
s’agit donc d’une simple bande de papier d’environ
Et
je vais essayer, en parcourant cette échelle, de vous donner un genre de « crash course » hyper résumé de morphologie évolutive des chordés,
les chordés étant la lignée des animaux qui ont développé une colonne vertébrale
avec une moelle épinière dedans et un cerveau au bout (en gros, les poissons,
amphibiens, reptiles, oiseaux, mammifères) contrairement aux invertébrés (mollusques,
étoile de mer, etc) qui n’en ont pas. Et je fais
ça toujours dans le but, je l’espère, de vous faire sentir un peu mieux la
longue histoire du cerveau humain qui, vous commencez peut-être à le pressentir,
est un peu plus longue que la seule histoire de l’hominisation…
Donc
si on regarde ce qui se passe à partir de 600 millions d’années, on
assiste à l’apparition d’organismes pluricellulaires,
i.e. contrairement aux bactéries qui étaient constituées que d’une seule cellule,
des organismes qui en contiennent plusieurs (il y en a jusqu’à 100 000
milliards dans le corps humain), d’où l’utilité d’avoir un système de communication
entre ces différentes cellules, ce qui sera le début des systèmes nerveux.
Je
vais vous monter ici, dans le thème sur le développement du cerveau humain
au cours d’une vie
(cliquer https://lecerveau.mcgill.ca/flash/i/i_09/i_09_cr/i_09_cr_dev/i_09_cr_dev.html )
le tout début d’un embryon humain après la fécondation, ce qui ressemble vraisemblablement
pas mal à l’évolution des premiers organismes pluricellulaires.
Donc
un amas de 2, 4, 8, 16, 32 cellules… qui a pu former éventuellement une sphère (on voit les cellules tout autour)… qui a pu ensuite se replier vers l’intérieur…
[descendre
sujet 2] On retrouve ici la sphère vue du côté de l’invagination en vert avec
en même temps une coupe transversale. On voit que la sphère s’allonge et quand
l’invagination se referme, cela forme un espèce de tube qu’on appel le tube neural et qui deviendra la moelle épinière.
(cliquez
https://lecerveau.mcgill.ca/flash/d/d_09/d_09_cr/d_09_cr_dev/d_09_cr_dev.html#3
)
Et
ensuite, ce qui va se passer, c’est que la partie antérieure de ce tube va
commencer à produire des renflements, d’abord 3 (prosen, mésen et rhombencéphale),
(cliquez
https://lecerveau.mcgill.ca/flash/i/i_09/i_09_cr/i_09_cr_dev/i_09_cr_dev.html#3
)
puis ces trois-là vont en se différencier en 5 renflements que l’on va retrouver jusque
dans l’anatomie du cerveau humain comme on l’a vu au début de la présentation.
Mais
revenons donc à il y a 500 millions d’années où la
vie explose en diversité avec l’apparition de l’embranchement des chordés
et du sous-embranchement des vertébrés. C’est la classe des poissons qui prospère la première, pendant
que sur la terre les premières plantes et les fougères commencent à apparaître.
À
quoi ressemble le cerveau des poissons ? À ceci...
http://trc.ucdavis.edu/mjguinan/apc100/modules/nervous/grosscns/brain9/brain.html
On
reconnaît d’abord la forme générale avec les différents renflements, avec
la partie la plus antérieure qu’on appelle le télencéphale ou le cortex,
(cliquez pour l’allumer et
faire remarquer qu’il n’est pas de taille importante),
les bulbes olfactifs qui en font partie, les lobes optiques
qui correspond au 3e renflement aussi appelé mésencéphale, le 4e
renflement ou métencéphale correspond au cervelet ici, et le bulbe rachidien
ou 5e renflement ou encore myélencéphale, suivi de la moelle épinière
qui va ensuite se poursuivre tout au long de la colonne vertébrale pour recevoir
les afférence sensoriel du corps et aussi envoyer les terminaisons nerveuses
des motoneurones aux muscles du corps.
Par
la suite, autour d’il y a environ 400 millions d’années, alors que
la végétation terrestre commence à être plus luxuriante et à offrir des ressources
intéressantes pour les animaux, les premières espèces d’amphibiens
apparaissent. Les amphibiens sont donc des organismes terrestres mais qui
dépendent encore du milieu aqueux pour leur reproduction. Des animaux, donc,
avec un système de branchies au stade larvaire
puis, à la métamorphose, des poumons ainsi
qu’une respiration cutanée toujours importante.
Voici
donc un cerveau d’amphibien, celui d’une
grenouille :
http://www.umanitoba.ca/science/biology/lab18/biolab18_4.html
On
reconnaît les mêmes structures que chez le poisson, avec le cortex (qui est
peut-être un peu plus allongé), les lobes optiques, le cervelet, la médulla,
la moelle épinière…
Toujours
il y a environ 3-400 millions d’années, les insectes colonisent peu
à peu tous les milieux. Dans la lignée qui a menée jusqu’à nous, on voit apparaître
les reptiles dont le cerveau devait ressembler
à peu près à ce cerveau de caïman
http://www.britannica.com/eb/art-70848
Les
bulbes olfactifs demeurent une structure importante chez les reptiles. Mais
on remarque que le cortex devient relativement plus important que les lobes
optiques.
Puis
pendant le règnes des dinosaures qui commence il y a environ 250 millions
d’années, arrivent enfin les premiers petits mammifères
qui sont d’abord ovipare, puis marsupiaux, puis finalement vers la fin de
l’ère des dinosaures il y a 65 millions d’années les mammifères à placenta
comme nous. Ou comme le rat dont on
voit le cerveau ici avec celui d’un alligator : http://www.neurobio.arizona.edu/282/Lectures2005/Brain%20structure/07_024.jpg
{kind=link}
pour montrer
qu’en même temps que se produit des changements au niveau macroscopique, par
exemple la diminution des bulbes olfactifs au profit du cortex, se produit
aussi des changement au niveau microscopique,
au niveau des neurones. Par exemple on voit ici une coupe au niveau du
cortex qui montre son épaississement chez les mammifères où l’on distingue
jusqu’à 6 couches alors qu’il n’y en n’a que 2 chez les reptiles. Donc en
plus de la surface corticale qui devient plus grande, il y a plus de couches
donc plus de neurones donc plus de possibilités de connexions.
C’est
donc seulement à partir d’il y a 65 millions d’années avec les niches
écologiques laissées vacantes par la disparition des dinosaures, que les mammifères
vont véritablement se développer et donner naissance aux primates et, il y a seulement 3-4 millions
d’années (certains disent 6-7), aux premiers représentants du genre Homo
dont nous faisons partie.
On
peut observer ici
(cliquer sur https://lecerveau.mcgill.ca/flash/i/i_05/i_05_cr/i_05_cr_her/i_05_cr_her.html
)
l’évolution
de la morphologie de quelques cerveaux de mammifères. Du rat
à l’homme en passant par le lapin,
le chat, le mouton et le chimpanzé.
Il
y a aussi un cerveau de dauphin
pour montrer que le cerveau humain est loin d’être le plus gros cerveau animal,
et que les gros animaux ont souvent de plus gros cerveau que nous car ils
ont une plus grande surface corporelle à innerver et plus de muscle à contrôler.
On les voit ici à gauche à l’échelle.
Ceci
dit, ce qui est beaucoup plus révélateur que la grosseur absolue du cerveau
c’est le poids
relatif du cerveau par rapport au poids du corps, et à ce niveau, les
êtres humains sont les champions. On remarque aussi la quantité de plus en
plus grande de circonvolutions
au niveau du cortex, ce qui indique une surface corticale de plus en plus
grande qui culmine chez l’humain avec nos
Observation
très importante : cette tendance à l’augmentation de la taille relative
du cerveau par rapport au corps ne montre pas une progression régulière chez
les primates, mais plutôt une courbe
avec une cassure spectaculaire, à partir d’environ 3-4 millions d’années.
Je
vais essayer de vous le dessiner au tableau un graphique toujours très « high-tech »
pour vous montrer l’ampleur de cette cassure (voir graphique ci-bas)
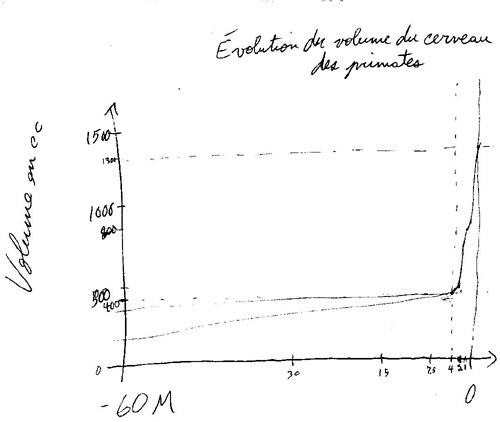
Temps
La
question qui se pose est évidemment qu’est-ce qui s’est donc passé durant
ces 3-4 derniers millions d’années dans la boîte crânienne de nos ancêtres
?
Une
première remarque qu’on peut faire ici, c’est que ces
3-4 millions d’années correspondent au paléolithique, i.e. en gros à l’évolution
du genre homo jusqu’à nous, et donc la période durant laquelle il y aurait
eu sélection de modules cognitifs selon les
principes de base de la psychologie évolutive.
Par
conséquent, on peut donc admettre qu’il
semble bel et bien s’être passé des choses importantes durant cette période
(en tout cas sur le critère de la surface corticale). Est-ce que cela justifie
toutes les formulations et les propositions classiques de la psychologie évolutive
? Ce n’est pas nécessairement le cas, et on va essayer de voir pourquoi dans
la suite de l’exposé…
Avec
la 4e partie qui porte sur les
IV- Richesses
et limites de la psychologie évolutive
Vous
connaissez sans doute mieux que moi les détails de la psychologie évolutive,
alors je vais me contenter de rappeler simplement l’idée
de base
(aller à https://lecerveau.mcgill.ca/flash/d/d_05/d_05_p/d_05_p_her/d_05_p_her.html#2
)
qui est que le système nerveux qui produit nos comportement individuels
est non seulement le fruit de notre histoire personnelle mais aussi, et surtout,
de l’histoire évolutive de notre espèce .
Et
que cette période est très longue, et ne s’est pas passé dans un environnement
comme le nôtre, une ville par exemple… En fait, près de 99% de l’histoire évolutive de Homos sapiens s’est
déroulée dans un environnement ressemblant à l’actuelle savane africaine. Durant toute cette période de 3-4 millions d’années, il semble que nos ancêtres vivaient en petits groupes
de chasseurs-cueilleurs nomades.
Et c’est seulement il y a 10 000 ans que certains ont commencé à devenir sédentaires
et à pratiquer l’agriculture.
L’idée
maîtresse de la psychologie évolutive est donc de considérer le cerveau humain
actuel comme un vaste ensemble « d’unités spécialisées » ou encore
de « modules adaptés
aux problèmes rencontrés par nos ancêtres chasseurs-cueilleurs »
dans leur environnement. Cette approche confère à la psychologie évolutive
une grande puissance explicative, comme vous le savez sans doute et comme
l’ont bien montré Luc et Pierre dans leur article « UN DÉFI POUR LA PSYCHOLOGIE ÉVOLUTIONNISTE » que j’ai lu avec intérêt.
Mais
avant de partir à la recherche de ces fameux modules, je n’ai pas
pu résister à la tentation de vous détailler un peu cette période qui correspond aux 3-4 derniers centimètres de ma
première bande de papier, pour la développer sur… une 2e grande bande d’environ
[sortir la deuxième bande
]
qui
est elle aussi à l’échelle, et qui résume de façon très générale les 4 derniers
millions d’années, avec l’époque des australopithèques, de Homo habilis,
de Homo erectus, Homo néanderthalis, etc.
jusqu’à l’homme de Homo sapiens sapiens ou Cro Magnon … (décrire ce que j’ai sur la bande).
Et
puis tant qu’à y être, pour aller jusqu’au bout de mon petit « trip »,
je me suis permis de détailler encore les derniers 2-3 centimètres… sur une
3e grande bande d’environ
[sortir la troisième bande
]
soit la période des 40 000 dernières années qui détaille
l’époque de Cro-Magnon, avec par exemple ici l’épisode des grottes de Lascaux,
puis l’apparition de l’agriculture il y a environ 10 000 ans, et finalement
les premières civilisations autour de 5 000 ans, l’Antiquité grecque,
Jésus-Christ il y a 2000 ans, l’empire Romain, Moyen Âge, Montaigne, etc… jusqu’à Darwin, Einstein et Freud !
Donc
je vais laisser ça affiché, comme ça on pourra y référer au besoin, et je
vais poursuivre en rappelant quelques
critiques qui ont été formulées à l’endroit de la psychologie évolutive.
Et
évidemment, je vais me servir du point de vue d’un neurobiologiste, celui
du Dr. Jaak Panksepp, qui est un spécialiste des émotions, et qui
a identifié :
1) Les
« sept péchés capitaux » de la psychologie évolutives pour un neurobiologiste
! Selon lui, la psychologie
évolutive pécherait entre autre pour les 7 raisons suivantes que j’ai regroupé
pour pouvoir le commenter un peu :
Péché
1) elle accorde trop d’importance au Pléistocène (donc
à l’hominisation) comme source des adaptations de l’esprit humain que l’on
constate aujourd’hui;
Péché
2) elle fait de l’anthropocentrisme en se concentrant
exclusivement sur l’être humain, délaissant la profonde continuité qui existe
en particulier dans les processus sous-corticaux
des mammifères;
Péché
5) elle attribue à des modules corticaux des mécanismes
émotifs dont les bases neuro-physiologiques sont
plutôt bien établies à des niveaux sous-corticaux
beaucoup plus anciens;
Ce
que je voudrais dire ici c’est qu’à la lumière de la morphologie évolutive
des chordés dont on a dressé tantôt un rapide portrait, ces deux critiques
me semblent assez bien fondée. En effet, il est clair que plusieurs des comportements
que les psychologues évolutionnistes ont tenté d’expliquer par des modules
corticaux font appel à des structrues sous-corticales que possédaient déjà tous les mammifères,
et parfois même les reptiles. Je pense à des comportements comme la jalousie,
qui font très certainement appel au système limbique mammalien et même à certains
noyaux hypothalamiques. Dans le cerveau humain, ces structures anciennes ont
bien entendu tissé au fil du temps des connexions avec le cortex, mais reste
que leur influence doit être tenu en compte dans une explication qui se veut
globale d’un phénomène comme la jalousie par exemple.
Péché
3) affirme que la psychologie évolutive serait trop fortement
adaptationniste,
ayant tendance à considérer tous les «modules» de la pensée humaine comme
des adaptations, alors que plusieurs facultés de l’esprit humain pourraient
tout aussi bien s’avérer être des exaptations, c’est-à-dire comme Stephen J. Gould l’a bien expliqué
avec son concept de « spandrels » emprunté à l’architecture, des structure qui existent
non pas parce qu’elles ont été sélectionné elles-même,
mais simplement parce qu’elle sont
des conséquences d’autres structures qui, elles, ont été sélectionnées.
L’exemple classique que donne Gould en architecture est le triangle qui se
forme lorsqu’on juxtapose une coupole sur 4 piliers : le seul fait de
juxtaposer ces 2 formes crée 4 triangles aux 4 coins, et même si les peintres
s’en servent dans les églises pour peindre de belles fresques, aucun architecte
n’a jamais voulu que ces triangles existent. Ils découlent simplement d’un
ensemble plus complexe, et tant qu’à être là, ils sont utilisés par les peintres.
Et comme la nature est une grande bricoleuse, je pense qu’il y a de fortes
chances pour qu’elle ait effectivement utilisé bien des exaptations
pour nous bricoler quelques facultés cognitives qui n’ont jamais été sélectionnées…
Cette
critique est assez bien résumée par le « requin-cornet »
de cette caricature : (cliquer sur https://lecerveau.mcgill.ca/flash/i/i_05/i_05_p/i_05_p_her/i_05_p_her.html,
puis aller tout en bas de la page):
Péché
4) serait qu’une trop grande importance est accordée à
la modularité du cerveau au détriment
des aires associatives à usage général, très importantes dans le cerveau
humain;
(cliquer sur https://lecerveau.mcgill.ca/flash/i/i_05/i_05_cr/i_05_cr_her/i_05_cr_her.html
, et descendre au milieu de la page)
En
effet, on constate aussi que, du rat à l’être humain, l’importance relative des aires sensorielles diminue au profit d’un cortex qu’on peut qualifier de manière
très générale d’associatif,
avec de plus en plus de neurones qui servent
à mettre en relation entre elles des régions du cortex. Il est vrai qu’on
a aussi découvert des fonctions particulières à plusieurs groupes de neurones
dans ces aires, mais il est probable que la fonction d’une proportion importante
des neurones de ces aires soit tout de même assez générale.
Enfin,
les deux derniers « péchés », que je me contenterai d’énoncer :
Péché
6) la psychologie évolutive abuse des métaphores pour
expliquer les mécanismes mentaux tout en ne se souciant pas assez de les confronter
aux données neuro-anatomiques existantes ;
Péché
7) elle est trop centrée sur une approche de représentation
et de calcul et pas assez incarnée dans le substrat organique.
Quelles
sont donc ces données recueillies depuis plusieurs décennies sur la neuro-anatomie
du cerveau non seulement de l’humain mais aussi des autres mammifères, et
qui nous permettrait d’essayer de cerner un peu ce que pourraient être les
modules ou les systèmes spécialisés de ce « substrat organique »
que constitue le cerveau ?
Le
titre de la dernière section m’a été inspiré des fameux livres « Où est
Charlie ?» (où il faut trouver un petit personnage parmi des dizaines d’autres
dans une page), une section, donc, qui s’intitule simplement :
2) Où sont les modules ?
Les dernières décennies de recherche en neurosciences
ont permis d’identifier d’abord des aires corticales et des noyaux sous-corticaux impliqués dans les fonctions sensori-motrices.
(aller à https://lecerveau.mcgill.ca/flash/a/a_06/a_06_cr/a_06_cr_mou/a_06_cr_mou.html
)
Pour la motricité, on a identifié par exemple un certain
nombre d’aires corticales, dont les fonctions se complexifient et deviennent
moins claires à mesure que l’on passe du cortex moteur primaire, au cortex
prémoteur, à l’aire
motrice supplémentaire, au cortex
préfrontal dorso-latéral et au cortex pariétal postérieur.
(descendre sujet 2)
De même, on connaît plusieurs noyaux sous-corticaux (i.e. sous le cortex) qu’on appel les ganglions de la base et qui jouent un
rôle aussi très important dans le mouvement. Plusieurs des voies qui relient
ces noyaux sont aussi connues et se révèlent d’une grande complexité. Sans
parler du cervelet, qui lui aussi
est grandement impliqué dans la coordination des mouvements, et dont les connexions
avec toutes les autres aires impliquées dans le mouvement sont riches et complexes.
(aller à https://lecerveau.mcgill.ca/flash/a/a_02/a_02_cr/a_02_cr_vis/a_02_cr_vis.html#2)
Du côté d’un système sensoriel comme la vision par exemple,
c’est la même chose. De la rétine, on connaît les premiers relais sous-corticaux dans le corps
genouillé latéral du thalamus, ou dans les collicules supérieurs. Ces relais envoient ensuite l’information aux aires visuelles primaires
(descendre sujet 3)
qui sont situées dans le lobe occipital ici. Différentes aires appelées V2, V3, V4, V5 / MT, etc, sont
spécialisées dans le traitement de différents aspects du stimulus visuel,
comme la couleur, le mouvement, les angles, etc. Aires qui sont aussi interconnectées
entre elles et définissent en gros deux
grandes voies, la voie ventrale
davantage impliquée dans la reconnaissance de la forme des objets et de leur
valeur affective; et la voie dorsale
qui servirait plus à guider en temps réel les actions que nous dirigeons vers
des objets.
On voit donc se dessiner un premier grand niveau de circuits interconnectés qui sont localisés
dans certains lobes corticaux, tout en pouvant en sortir, et qui sont associés
à une fonction générale, sensorielle ou motrice. Peut-on parler de modules
à ce niveau au sens où les psychologues évolutionnistes en parlent ? Je ne
sais pas, mais j’en doute… Ne serait-ce que parce que, comme on l’a vu, ces
circuits font à la fois intervenir le cortex mais aussi bon nombre de structures
sous-corticales beaucoup plus anciennes.
Justement, dans le cas des structures plus anciennes
qui facilitent l’expérimentation animale puisqu’elles sont présentes chez
le rat par exemple, on a pu investiguer davantage. En effet, si l’on prend
l’étude d’une émotion comme la peur
par exemple,
(aller à https://lecerveau.mcgill.ca/flash/d/d_04/d_04_cr/d_04_cr_peu/d_04_cr_peu.htm
)
on a tout d’abord pu identifier les principales structures
impliquées, comme dans le cas de
Mais encore ici
(descendre au sujet 2), d’autres structures comme le thalamus ou les différents
cortex sensoriels sont impliqués, et on a pu montrer qu’il existe ce qu’on
appelle une route courte qui va
directement du thalamus à l’amygdale, où une réponse émotionnelle peut être
donnée avant même que la source potentiellement dangereuse n’ait été complètement
discriminée par une autre voie qu’on appelle la « route longue » du cortex. Un exemple
de ça c’est le pied que vous retirez rapidement à
la vue d’une forme allongée dans les herbes hautes, avant de vous apercevoir
que ce n’était pas un serpent mais simplement un vieux boyau d’arrosage…
(aller à https://lecerveau.mcgill.ca/flash/d/d_04/d_04_cl/d_04_cl_peu/d_04_cl_peu.htm
)
Mais ici on peut aller encore plus loin en distinguant
plusieurs amas neuronaux distincts
à l’intérieur même de l’amygdale, dont le noyau latéral, qui reçoit la majorité de l’information afférente,
et le noyau central, qui est plutôt
le point de départ des outputs.
(cliquer sur https://lecerveau.mcgill.ca/flash/i/i_04/i_04_cl/i_04_cl_peu/i_04_cl_peu.htm
)
Et, bien sûr, on commence à identifier des voies spécifiques
qui relient chacun de ces noyaux. Commence-t-on à discerner dans ces sous-régions
plus limitées des cibles
plus envisageables pour un processus de sélection adaptative ? Peut-être.
Sauf que dans cet exemple-ci, je vous rappelle qu’on est dans l’amygdale,
donc dans le système limbique, et non dans le cortex…
Revenons-y donc justement, au cortex. Parce que différentes
techniques de marquage et d’électrophysiologie ont permis la reconnaissance de spécificités fonctionnelles
à des niveaux encore plus fins. Pour vous donner juste un exemple de ça,
je vais revenir au cortex visuel...
(aller à https://lecerveau.mcgill.ca/flash/a/a_02/a_02_cl/a_02_cl_vis/a_02_cl_vis.html#3
)
…ou l’on a pu discerner une organisation neuronale en
colonnes. C’est qu’en
plus de l’organisation en 6 couches horizontales caractéristique du cortex,
on s’est aperçu que tous les neurones qui se trouvent dans
une même petite colonne verticale répondent
tous à une même caractéristique d’un point donné du champ visuel (ici,
l’orientation oblique d’une ligne par exemple). Les colonnes forment donc
une unité fonctionnelle perpendiculaire à la surface corticale. Et si on fait
des enregistrement sur une trajectoire parallèle à la surface du cortex (donc
dans le sens horizontal), on va traverser plusieurs colonnes et donc rencontrer
autant d’orientation possible pour notre ligne.
Chaque colonne a un
diamètre d’environ 300 mm, s'étend sur toute l'épaisseur
du cortex, soit environ 3mm et contient quelques milliers de neurones. Cette
organisation en colonnes, elle a été retrouvée un peu partout dans le cortex.
Les colonnes possèdent une connectivité
de proximité avec les colonnes voisines, mais aussi à grande distance (aire à aire ou hémisphère à hémisphère), par le
biais d'axones myélinisés.
Alors est-ce que les colonnes pourraient être des « modules
spécialisés » au sens de la psychologie évolutive, ou au contraire
simplement des unités générales de traitement de l’information
du cortex ? Personnellement, je n’ai pas les compétences pour me prononcer
là-dessus…
Ceci dit, le fait est qu’on peut, dans le système visuel, identifier certaines caractéristiques
dont la fonction laisse effectivement
penser qu’il y a pu y avoir une sélection
pour un type de stimuli particulièrement
significatif pour nous. Et je vais terminer en
vous donnant deux exemples de ça.
Dans le premier exemple, il s’agit toutefois d’un stimulus
qui a probablement été significatif non seulement pour nos ancêtres humains,
mais pour les autres animaux aussi.
Il s’agit de notre compréhension implicite, inconsciente
et instantanée de l’effet de perspective. On en a un exemple ici :
http://decision.csl.uiuc.edu/~giridhar/Photos/campus/slides/perspective.html
avec les lignes convergentes vers un point de fuite qui
nous font ressentir tout de suite l’éloignement en allant vers ce point de
fuite.
Ce phénomène visuel suggère l’existence dans notre cerveau
des circuits spécialisés pour déceler rapidement et automatiquement ce type
de stimulus. Et ces circuits, solidement établis dans notre cerveau peut-être
dès la naissance, seraient à la base de plusieurs de nos « illusions d’optique ».
(aller
à https://lecerveau.mcgill.ca/flash/i/i_05/i_05_p/i_05_p_her/i_05_p_her.html
)
Par
exemple, les deux rectangles blancs ici sont bel et bien de la même longueur
bien qu’on jurerait que celui du haut est plus long. L’explication est la
suivante : pour le cerveau, plus quelque chose est près du point de convergence
de plusieurs lignes, plus celui-ci le considère comme étant loin, car le point
de fuite vers l’horizon est un phénomène optique qui de tout temps nous a
donné une idée de
Et
même avec toutes ces explications, c’est très difficile de la percevoir comme
étant de la même longueur, ce qui montre que de tels « modules »
spécialisés sont très efficaces et fonctionnent tout à fait inconsciemment. Quant
à savoir quels sont les circuits sous-jacents qui en sont la cause, personnellement
je l’ignore …
On
a ici (aller à https://lecerveau.mcgill.ca/flash/i/i_02/i_02_p/i_02_p_vis/i_02_p_vis.html
)
un autre exemple de cette illusion, avec la preuve en
plus, pour les sceptiques…
Mon
autre exemple est plus spécifique aux humains et concerne la reconnaissance
des visages. Il semble en effet qu’on ait une prédisposition toute particulière
pour reconnaître les visages. En fait, on est souvent prompt à en voir partout.
Comme
le montre bien l’article précédent, on voit souvent des visages dans les nuages,
dans une formation rocheuse sur la planète mars, dans la fumée des tours en
feu du World Trade Center, et même dans un grill
cheese qui s’est vendu 28 000$ sur E-Bay il n’y a pas longtemps parce des chrétiens ont dit qu’il
s’agissait du visage de
L’importance
de la reconnaissance des visages pour
comprendre les expressions et les intentions des autres constitue certainement
un stimulus hautement significatif pour les humains, et notre faculté d’être
hypersensible à tout ce qui ressemble à un visage est cohérente avec l’idée
de modules spécialisés pour les détecter. On peut aussi le dire autrement :
le coût, en terme de survie, d’échouer à reconnaître
un visage est plus élevé que le coût de percevoir un visage là où il n’y en
a pas…
Or
il existe des patients qui, à la suite d’un accident cérébro-vasculaire,
peuvent reconnaître tous les objets SAUF
les visages ! On nomme ce syndrome rare
(aller
à https://lecerveau.mcgill.ca/flash/a/a_01/a_01_cr/a_01_cr_ana/a_01_cr_ana.html
))
qui étaient fortement activées par les visages contrairement
aux autres objets. Ils ont ensuite pousser leur investigation avec des techniques
d’enregistrement intracellulaire dans ces régions du cerveau du macaque, et
les résultats ont été sans équivoque : ces cellules répondaient
uniquement aux visages ou aux stimuli qui ressemblaient à des visages
et pas aux autres. Ces résultats ont d’ailleurs été publiés en octobre
dernier dans
Alors
voilà, on pourrait dire qu’on a enfin
ici quelque chose qui ressemble à un
module spécialisé issu probablement d’un processus de sélection. Mais
malheureusement, encore une fois, ce n’est pas aussi simple que ça…
Car
un autre phénomène, très étudié par un neurobiologiste de l’université de
Boston, le Dr. Takeo Watanabe, pourrait aussi contribuer
à notre grande facilité à percevoir les visages. Ses travaux, qui portent
sur l’apprentissage, montrent qu’après
avoir été bombardé par un certain type de stimulus, le cerveau tend à continuer
de percevoir ce stimulus même s’il n’est plus présent.
Voilà
pourquoi ce chercheur pense que le
très grand nombre de visages que nous voyons chaque jour pourrait aussi être
en partie responsable pour notre prédisposition à voir des visages partout.
Autrement dit, plus on en voit, plus on est porté à en voir…
Mais
alors, on peut aussi se demander si cette capacité de pouvoir être sensibilisé
à un certain type de stimulus pour le reconnaître plus facilement ne pourrait
pas elle aussi nous avoir procuré un avantage adaptatif
? (comme le fait
de pouvoir rapidement reconnaître un serpent une fois qu’on en a vu quelques
uns par exemple) et que donc, cette
capacité aurait pu constituer elle aussi un « module spécialisé »
hérité de nos ancêtres ? Ce n’est pas impossible…
Donc à travers ces
deux exemples de la perspective et de
la reconnaissance des visages, j’ai surtout voulu attirer l’attention
sur le fait que la nature et l’évolution sont de
grand bricoleurs, des bricoleurs empiriques, bref quelque chose d’assez
loin des lois universelles et des concepts généraux qui veulent avoir un pouvoir
explicatif global et absolu comme c’est le cas de la « modularité cérébrale »
sélectionnée quand on ne s’en remet qu’à elle seule.
Et
je terminerai en vous montrant des images d’une expérience d’imagerie cérébrale
à laquelle j’ai participé comme « cobaye » il y a quelques années
à l’Institut Neurologique de Montréal…
(aller
à https://lecerveau.mcgill.ca/flash/capsules/experience_jaune04.htm
)
Et
on peut voir ici dans mon cerveau (et vous aller comprendre pourquoi je prends
mon cas particulier très vite) qu’il y a donc certaines
aires qui s’activent spécifiquement lorsqu’on me demandait d’effectuer certaines
tâches, comme par exemple ici :
1)
de générer des noms à la suite d’une description auditive d’un objet (on voit
l’activité
principale a lieu dans l’hémisphère
gauche, dans les aires associées à la compréhension et à la génération
du langage.
2)
ici durant
la tâche faisant intervenir la mémoire de travail : l’activité principale a
lieu dans le cortex préfrontal.
3)
ici sur une coupe sagittale durant une tâche d’imitation avec la main droite: l’activité
principale a lieu dans le cortex visuel et l’activité
secondaire dans le cervelet (coordination motrice).
4) ici durant la même tâche mais sur une coupe coronale
: l’activité principale a lieu dans le cortex moteur de l’hémisphère gauche et l’activité
secondaire dans les deux cortex visuels ventraux.
Ceci
dit, ça c’était pour mon cerveau, mais il ne faut jamais oublier que chaque cerveau est unique et qu’il existe
une très grande variabilité inter-individuelle
dans l’activation cérébrale pour une même tâche, comme on le voit ici (remonter un peu dans la capsule) où l’on
a les réponses de 5 individus différents pour deux tâches distinctes.
Bref, comme le montrent les résultats d'imagerie cérébrale,
on peut dire en guise de conclusion qu’il semble que le traitement de l'information
par le cerveau consiste en un dialogue ordonné entre des aires ou des « modules »
mais qui sont activés de façon simultanée
en des régions parfois distantes du cerveau et qu’il demeure très difficile
d'identifier les interactions entre ces modules, et encore plus, leur origine…
FIN.